They make up about over 90mm of all animals on Earth, yet most people do not acknowledge their presences. Again everyone knows the cute and lovable Koala bear, believe all snakes are deadly, a slippery fish but when it comes to arthropods they are left in the dark. Oh no not those creepy crawlies, bang splat got it. They play a major role in maintaining ecosystems as pollinators, recyclers of nutrients, scavengers and food for other animals yet most people detest them, have phobias at the sight of them, and cannot name even a hand full of them. Their lives remain a secret to all but the Arachnologist, Carcinologist, Entomologist and Myriapodologists.
Despite being the most abundant species on the planet very little is known about the complex relationships that exist between artropods, flora and their biodiversity influences they have even on the basic fundamenltals of an ecosystem. Arthropod driven functions, like herbivory and pest-control, with their direct and indirect associations among other arthropods flora and fauna are still basically in the initial stages of science. Very few studies and none to my knowledge have been undertaken on the affects of arthropods on depleting natural habitats, global warming and the extinction of so many plant communities. Most studies in the past have been restricted to the elimination of species, which compete in consuming human crop production.
The higher a plant species richness in a habitat, the greater the diversity and the greater the complexities of arthropod communities yet we have failed to note in any detail the intricacies of these vital webs. All natural ecosystem’s would have substantially more herbivore and carnivore diversity, proportionally as the richness and health of an ecosystem increases, but to what extent is still relatively unknown. When I mentioned to Alex Floyd that “I would like to describe with my camera the connection between plants and their associated arthropods in the district.” Alex replied “You will open up your camera to a gigantic can of worms with no starting point and no conclusion. Good luck.”
Identifying: Arthropoda
Arthropods are the most numerous “phylum” of the animal kingdom with about the 95mm of all known species belong to this “phylum”. Arthropods are easily identified because they are invertebrates with articulate legs. These two characteristics give them their Phylum name.
Usually they have two compound eyes; formed by many eyes called ommatidia as in flies and or several ocelli, simple eyes as in the 8 eyes of a spider.


Most of them are oviparous in that they lay eggs though some members are viviparous bearing live young while others are ovoviviparous where the incubation of the eggs takes place internally and the eggs hatch as they are being laid.
Generally arthropods are subject to the metamorphosis process. The other terrestrial invertebrate animals belong to the Phyla Anellida known as worms and the Mollusca class of Gastropoda.
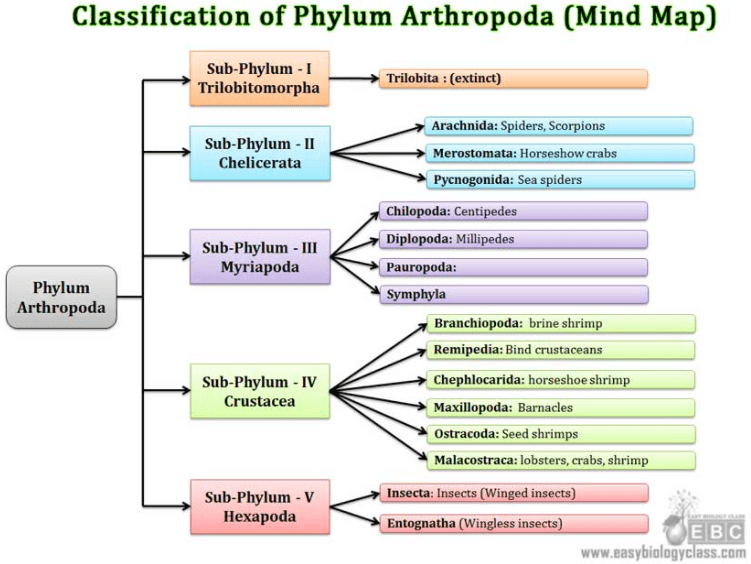
Arachnida
Generally the Arachnid’s body is divided in two sections the cephalothorax with the thorax and head being fused together and abdomen. With the Opiliones and Acari specie the sections are fused together. They have neither wings nor antennae.
They have four pairs of legs with two pairs of appendages known as the chelicerae and pedipalps which are usually specialized to catch and kill prey. Most specie are carnivorous.
Generally respiration is pulmonary. Arachnids have only simple eyes and are oviparous or viviparous.

Crustacea
Crustacean literally means dermoskeleton. The external body layer is made of chitin and is the same material that fingernail and toenails are made. It is strong and generally covered by carapace a calcareous material. Most of them live in the sea, but some live in rivers, lakes, ponds while a few are terrestrial species. Their body is divided in the thorax with head being fused together and abdomen.
They have two pairs of antennae and are oviparous. Generally the respirator mechanism is brachial.
Some orders of this class are Cladocera where the abdomen is not segmented and the bodies are flat. The Isopoda are without carapace and the legs are similar. Most of the terrestrial Arthropods in this order include those of the Armadillidiidae which can roll into balls and those in the Porcellionidae family which can’t roll into ball. The Decapoda which still have carapace have the first three pairs of legs fitted to catch and triturate food.

Insecta
insects are the most numerous class within the Arthropods. Insects are easily indentified because the adults have three pairs of legs. It is the only class which has wings or wing remnants in the adult stage.

Myriapods
The Myriapods and insects share several derived traits representing adaptations to land: tracheal systems, predominantly uniramous (unbranched) limbs and excretory Malpighian tubules opening into the hindgut.
Myriapoda body is divided in two sections: head and trunk, in fact thorax is fused with abdomen. The body is divided into many sections with one or two pairs of legs on each section. The antennae are long and usually filiform.
The respiration is either cutaneous or tracheal. They are oviparous and most are nocturnal in habitat.
Myriapods have separate sexes (gonochoristic), and sperm transfer generally involves the exchange of one or more spermatophores via specialized appendages.
Myriapoda comprise four classes of terrestrial arthropods which are the Chilopoda (centipedes), Diplopoda (millipedes), Symphyla and Pauropoda.


Symphyla – https://en.wikipedia.org/wiki/Symphyla
The oldest unequivocal myriapod fossil is of the millipede Pneumodesmus newmani, from the late Silurian which is around 428 million years ago). Pneumodesmus newmani is also important as it is the earliest known terrestrial animal and probably started colonizing land before insects. (Lower photos Rowland Shelley & Paul Marek (March 1, 2005). “Millipede Fossils”. East Carolina University)
Arachnids
Characteristics of Arachnids: The respiratory system of Arachnids is typical of smaller type terrestrial organisms and differ from there ocean cousins the crustaceans. The internal respiratory surfaces take the form of tracheae or modification of the book gill into a book lung with an internal series of vascular lamellae used for gas exchange with the air. While the tracheae are often individual systems of tubes, similar to those in insects. The Ricnuleids, Pseudoscorpions and some spiders have sieve tracheae, where several tubes rising to form a bundle from a small chamber connected to the spiracle. This type of tracheal system has almost certainly evolved from the book lungs, and indicates that the tracheae of arachnids are not a mixture of genes shared with insects.
Book lungs are not related to the lungs of modern terrestrial vertebrates. The name describes the appearance of the breathing organ. The appearance is that of pages of a book and has alternating air pockets and tissue filled with hemolymph. The number of pages or plates varies from just one pair in most spiders to four pairs in scorpions. The plates maximize the surface exposed to the air and thus maximize the amount of gas exchanged between the organism its environment. In most species, no motion of the plates is required to facilitate this kind of respiration. Book lungs are present in scorpions, Tetrapulmonata, whip scorpions, Schizomida, Ambypygi, and spiders.
The book lungs are absent in the microwhip scorpions, harvestmen, Acarina, Pseudoscorpions, Ricinulei and sun spiders.
Arachnids have a sexual reproductive system. Special sensory organs, and water conservation is enhanced by efficient excretory structures as well as a waxy layer covering the cuticle.
The excretory glands of arachnids include up to four pairs of coxal glands along the side of the prosoma, and one or two pairs of Malpighian tubules, emptying into the gut. Many arachnids have only one or the other type of excretory gland, although several do have both. The primary nitrogenous waste product in arachnids is guanine.
Arachnid blood is variable in composition, depending on the type of respiration system employed. Those Arachnids which have an efficient tracheal system do not need to transport oxygen in the blood, and may have a reduced circulatory system. In scorpions and some spiders the blood contains a copper based haemocyanin, which has a similar function to iron based haemoglobin, in vertebrates. The heart is located in the forward part of the abdomen, and may or may not be segmented. Some mites have no heart at all.
Body structures
Almost all adult arachnids have eight legs which makes them easy to recognize from their insect cousins which have just 6 legs. Arachnids also have two pairs of specialized appendages that are adapted for feeding, defense, and sensory perception. The first pair is the chelicerae which are used in feeding and defense. The second pair is the pedipalps which are used for feeding, locomotion, and reproductive functions. The pedipalps in Solifugae are more leg like, so that these animals appear to have ten legs. The larvae of mites and Ricinulei have only six legs with the fourth pair usually appearing when they moult into nymphs. However, mites are variable as there are mites with eight legs while some adult species have just four legs with a rare few possessing 6 legs.
The 8 leg appendages are modified for more efficient locomotion on land though some still retain the sideways and reverse characteristics of crabs. Most arachnids lack extensor muscles in the distal joints of their appendages. Spiders and whipscorpions extend their limbs hydraulically using the pressure from their hemolymph. Solifuges and some harvestmen extend their knees by the use of highly elastic thickenings in the joint cuticle. Scorpions, pseudoscorpions and some harvestmen have evolved muscles that extend two leg joints on the femur-patella and patella-tibia joints. The equivalent joints of the pedipalps of scorpions are extended by elastic recoil.
Arachnids do not have antennae and are wingless.
Their body is organized into two tagmata, called the prosoma, or cephalothorax, and the opisthosoma, or abdomen. The arachnid’s body is divided into two parts. The anterior section is known as the cephalothorax which is the combination or fusion of the cephalon (head) and the thorax (mid section) and is usually covered by a single, unsegmented carapace. The rear section or abdomen is segmented in the more primitive genre like the scorpion specie and is completely fused in more modern genre like the Acari specie. It is typically divided into a preabdomen and postabdomen.
The cephalothorax contains the sense organs, mouthparts, specialist limbs and limbs in pairs. The first pair of limbs is the chelicerae which usually form pincers or poison fangs, and the second pair is the pedipalps and may serve as pincers, feelers, or rudimentary to advanced legs. The other limb pairs, generally four, are used for mobility.
The posterior part of the abdomen bears the genital opening and other structures. It is usually equipped with modified gills called book lungs.
Argiope pulchella/Argiope aetherea (stabilimenta is very poor for A. aetherea but suits A. pulchella but the latter is not found in Australia?)
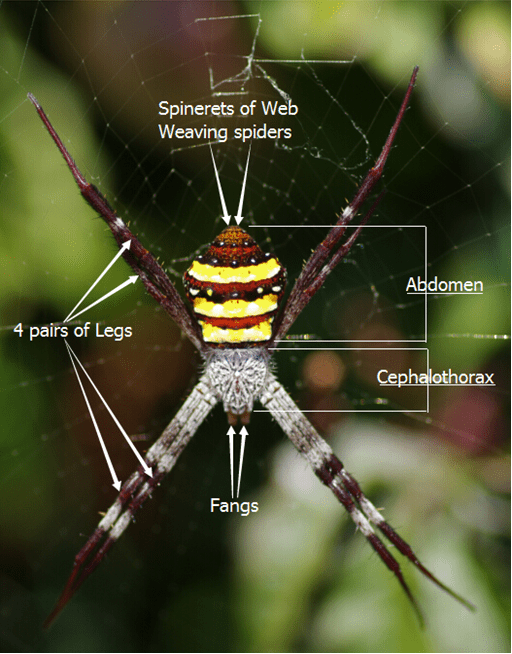
Basic characteristics of Arachnids include 4 pairs of legs, a body divided into two tagmata – the cephalothorax and abdomen and Fangs. The spinnerets for web spinning Spiders are shown.
Arachnids have an exoskeleton, and they also have an internal structure of cartilage like tissue, called the endosternite, to which certain muscle groups are attached. The endosternite has become calcified in some Opiliones.
Ecology Juveniles of some spiders in the families Anyphaenidae,
Corinnidae, Clubionidae, Thomisidae and Salticidae feed on plant nectar. Laboratory studies show that they do so deliberately and over extended periods, and periodically clean themselves while feeding. These spiders also prefer sugar solutions to plain water, which indicates that they are seeking nutrients. Since many spiders are nocturnal, the extent of nectar consumption by spiders may have been underestimated in the past. Nectar contains amino acids, lipids vitamins and minerals in addition to sugars and studies have shown that some spider species actually live longer when nectar is available. Nectar diets avoid the risks of struggles with prey, reduces the costs of producing venom, silk and digestive enzymes.
Various species are known to feed on dead arthropods (scavenging), web silk, and their own shed exoskeletons. Pollen caught in webs is also eaten, and studies have shown that young spiders have better chances of survival if they fed pollen. In captivity, several spider species are also known to feed on bananas, marmalade, milk, egg yolk and sausages. Jackson, R. R.; Pollard, Simon D.; Nelson, Ximena J.; Edwards, G. B.; Barrion, Alberto T. (2001).
Life Cycles Most arachnids are solitary except at the time of mating. Arachnids may have one or two gonads, which are located in the abdomen. The genital opening is usually located on the underside of the second abdominal segment.
Male spiders do not produce ready made spermatophores; packages of sperm, but spin small sperm webs on to which they ejaculate on and then transfer the sperm to syringe like structures on the tips of their pedipalps. When a male detects signs of a female nearby the rigors and dangers of passing his package to the female commences. He checks whether she is ready to mate; for example in species that produce webs or “safety ropes”, the male can identify the species and sex of these objects by “smell”.
Male spiders usually have rather elaborate courtship rituals to prevent the large females from consuming him prior fertilization; which is the norm, except where the male is much smaller than the female. In web weaving species, precise patterns of vibrations on the web are a major part of the rituals, while patterns of touches on the female’s body are important in most spiders that actively hunt. Gestures and dances by the male are important for jumping spiders, which have excellent eyesight. If courtship is successful, the male injects his sperm from his pedipalps into the female’s genital opening, known as the epigyne, on the underside of her abdomen. Female’s reproductive tracts vary from simple tubes to systems that include seminal receptacles where the females are able to store sperm and release it when she requires it. The advantage of sperm receptacles is that she only needs one mating for several broods or she can mix the sperm of several males giving her offspring greater chances of receiving the sperm from the best male. Of course the downside is if she is the hunted she may not get the opportunity to use the sperm.
In over 60mm of cases the female of the Australian redback spider kills and eats the male after it inserts its second palp into the female’s genital opening; in fact the males cooperate by trying to impale themselves on the female’s fangs. Observation shows that most male red back spiders never get an opportunity to mate, and the “lucky” ones increase the likely number of offspring by ensuring that the females are well fed. Most males in most species survive a few mating despite have shorter life expectancies than the females.
Female spiders lay up to 3,000 eggs in one or more silk egg sacs, which maintain a fairly constant humidity level. In some species the females die shortly afterwards while females in most species protect the sacs by attaching them to their webs, hiding them in nests, carrying them in the chelicerae or attaching them to the spinnerets.
Baby spiders hatch as spiderlings similar in all respects to their parents. The wolf spider’s brood clings to rough bristles on the mother’s back. The females of some species respond to the “begging” behaviour of their young by giving them their freshly killed prey, while others supply a regurgitated food source.
Like other arthropods, spiders have to moult to grow as the cuticle is unable to stretch. This is the signal and ideal time, in some species for the males, to mate with newly moulted females which are usually weakened to an extent that they are not a hazard to the males.
Scorpions are either ovoviviparous or viviparous, depending on species so virtually all the young are live born.
Most spiders live for only one to two years, although some tarantulas
have been known to survive in captivity for over 20 years.
Classification: If I wasn’t an Arachnophobiac before I certainly ain’t now as I grew to have a great affection for them following writing this section. From an early childhood; I have always had an fascination with spiders, now it has turned to a love that I may not be able to do these beautiful misunderstood organisms justice. Arachnids constitute the class Arachnida, in the phylum Arthropoda. If I have erred in any section please let me no so I can have it corrected.
The class is divided into 11 orders: the Acari or Acarina (mites and ticks), Amblypygi (tailless whipscorpions), Araneae (spiders), Opiliones (daddy longlegs), Palpigradi (palpigrades), Pseudoscorpiones (false scorpions), Ricinulei (ricinuleids), Schizomida (micro whipscorpions), Scorpionida (true scorpions), Solpugida (windscorpions) and Uropygi (whip scorpions).
Acari or Aracina: (Mites & Ticks)
Their bodies are not segmented (newly hatched young have 2 or 3 pairs of legs. They are very small.
The Argasidae or Bird lice without a dorsal shield are parasites of avian or birds.
Ixodidae or ticks have small dorsal shields and parasitize mammalian.
The Pyrogliphidae or dust mites are often found on birds while the Tretranichidae or clover mites often infest trees, vegetation and buildings.
Its fossil history goes back to the Devonian period, although there is also a questionable Ordovician record. The Devonian period was the time frame in which certain species of animals developed legs. In most modern treatments, the Acari is considered a subclass of Arachnida and is composed of two or three orders or super orders: Acariformes, Parasitiformes, and Opilioacariformes. Most acarines are minute to small and are less than 1mm in length though some of the Australian Ticks measure 10mm to 20mm in length when full gorged.
Only the slightest traces of primary segmentation remain in mites with the prosoma and opisthosoma being fused while a section of the flexible cuticle; the cirumcapitular furrow separates the chelicerae and pedipalps from the rest of the body. This anterior body region is called the gnathosoma (or capitulum) and is also found in the Ricinulei. The remainder of the body is called the idiosoma and is unique to mites. Most adult mites have four pairs of legs however a small number possess 2 or 3 pairs. Larval and prelarval stages have a maximum of three pairs of legs.
Acarines live in practically every habitat which includes aquatic and terrestrial species. They out number the other arthropods in the soil organic matter and detritus matter. Many are parasitic and they affect both vertebrates and invertebrates. Most Acarine specie is parasitic while the remainder is generally predaceous feeding on other Arthropods. A few are detritivores that help to break down forest litter mainly dead organic matter such as skin cells while a smaller number are herbivores and may damage crops. Damage to crops is perhaps the most costly economic effect of mites, especially among the spider mites and their relatives (Tetranychoidea), earth mites (Penthaleidae), thread footed mites (Tarsonemidae) and the gall and rust mites (Eriophyoidea).
Mites on humans often cause allergens and the stimulation of asthma in people affected by the respiratory disease. Predatory mites are used in the biological control of Phytoseiidae specie while various herbivorous mites are now being deployed to attack certain weeds is gaining acceptance.
Some of the parasitic ticks affect humans and other mammals, causing damage by their feeding, and can even be vectors of diseases such as scrub typhus (Rickettsial australis is transmitted by Haemaphysalis longicornis) and rickettsial pox (Rickettsial akari is mainly found in North America & Africa) while Lyssivirus have hypothesized is spread by Ixodes holocyclus (Refer Chapter 29 Human Wildlife Conflict).
Left Australian paralysis tick, Ixodes holocyclus. Note the fore and hind legs are a deeper colour than the central pair. The body shield is trapezeum. – unknown
Right Australian bush tick, Haemaphysalis longicornis. Note the legs are the same colour. The body shield is ovate. – wikipedia.org/wiki/Haemaphysalis_longicornis

Araneae: (Spiders)
Their bodies are divided into two sections, the cephalothorax and abdomen. All spiders produce silk, a thin, strong protein strand extruded by the spider from spinnerets most commonly found on the end of the abdomen.
Some of the most common families of spiders are:
Agelenidae or funnel web spiders which build funnel shaped webs ending in a tube which is their shelter and have long spinnerets. There are 40 described species in two genres (Hadronyche and Atrax) all from Australia. They are medium to large spiders, varying from 10mm to 50mm in body length. The males are more lightly built than their heavier built female counterparts. The body colour varies from black to brown with the carapace covering the front part of the body being sparsely haired and glossy. The lateral pair of spinnerets at the end of the abdomen is longer and easily visible in the species of Atrax and shorter in comparison in the Hadronyche species.
Not all species are dangerous, but several are renowned for their fast reacting possibly fatal venom. The male of Atrax robustus, the Sydney Funnel web Spider, is responsible for the 13 recorded death and many of the bites that require serious medical attention with the administration of antivenin. This remarkable spider has become a part of Sydney’s folklore and, although no deaths have been recorded since the introduction of an antivenin in 1981. It still retains great respect as the most venomous spider in the world.
Funnel web spiders make a burrow in moist, cool, sheltered habitats usually under rocks, in and under rotting logs and crevices. They can however be in the open adjacent to paths and other obstacles often utilizing natural holes and crevices. The preferred soils are sands and sandy loams. Tree dwellers prefer rot and borer holes in rough barked trees. In gardens, they prefer rockeries and dense shrubberies, and are rarely found in more open situations like lawns. The most characteristic sign of a Funnel web’s burrow is the irregular silk trip lines that radiate out from the burrow’s entrance of most species. These trip lines alert the spider to possible prey, mates or danger.
Funnel-webs are very vulnerable to drying out, so high humidity is more favourable to activity outside the burrow than dry conditions. Like most ground dwelling Arachnids they are nocturnal.
http://australianmuseum.net.au/Funnel-web-Spiders-group#sthash.V8E2nY0D.dpuf

Orangeade
Orb weavers are also known as garden spiders, often build beautiful large spiral webs. Araneae, or spiders, are the most familiar of the arachnids in Australia. There are around 40,000 species described world wide. Many species use it to trap insects in webs, although there are many species that hunt freely. Silk can be used to aid in climbing, form smooth walls for burrows, build egg sacs, wrap prey, temporarily hold sperm, and even fly, among other applications.
Left Golden orb Nephila edulis on web between two mangroves, Avicinnea marina. Right Unknown spider web at sunrise -andi Mellis

Ereiophora tranmarine different colour variations on Seringea denticulata leaf. – andi Mellis

All spiders except those in the families Uloboridae and Holarchaeidae, are in the suborder Mesothelae and have about 350 species. All are capable of injecting venom to protect themselves or to kill and to liquefy or aid in the digestion of prey. Only about 200 species, however, have bites that can pose health problems to humans. Many larger species’ bites may be painful, but do not produce lasting health concerns.
Spiders are found all over the world, from the tropics to the Arctic, while the water spiders live on or underwater in silken domes that they supply with air, and on the tops of the highest mountains.
Clubionidae
Sac spiders build a tubular sac which opens at both ends. They are found throughout Australia usually from moist forests to woodlands and grassed open woodlands. Slender Sac Spiders make small cylindrical or ovoid silk retreat sacs. They have slender bodies, large jaws and long, thin legs but not filiform. The males are much more slender than the females. Most are shades of cream, brown or yellow, with a darker stripe/s along the upper abdomen. The fangs of the male spiders are particularly enlarged. They measure 9mm to 16mm in length.
Slender Sac Spiders are common hunters on foliage in bush and garden. They make their retreat sacs in folded leaves or grass blades, but sometimes enter houses and buildings.
The Stout Sac Spiders may be found on house walls and fences but are most commonly found under loose bark and amongst leaf litter. Their sheet like sac retreats is often found beneath the bark of eucalyptus species.

Linyphiidae
These spiders build a web with a shape of a sheet or a dome. These spiders are closely related to the Clubionidae spiders and include around 400 species in about 30 genres worldwide. The largest genus currently recognized belong to the family Cheiracanthium, which used to be placed in the family of Clubionidae. The genus is quite diverse in Africa with a few in Asia and Australia. The family would be totally obscure if it were not for the fact that members of Cheiracanthium are documented as beneficial predators in agricultural grain fields.
The Lycosidae or wolf spiders do not build webs; the fourth pair of legs is the longest pair. The wolf spiders are very common and often form large communities in open paddocks and open woodlands from the coast to high in the mountains in Australia. Their presence is noticed by the holes and the beautifully fitted lid coverings of some other specie.
Wolf spiders carry their silk egg sacs by attaching them to their spinnerets (Pisauridae species carry their egg sacs with their chelicerae and pedipalps). Two of the Wolf spider’s eight eyes are noticeably large and prominent. They have excellent eye sight.
The percentage of spiders found in thistle cover or annual weeds is an indication that the spiders favour ground that has been recently disturbed and is going through rapid change in quality. This would indicate that the wolf spiders may benefit from habitat fragmentation, changing agricultural practices and wildlife corridors.
Opiliones or harvestmen are closely related to the Pholcidae but have their cephalothorax and abdomen fused or partially together and have long, filiform legs which bend above the body. Most species measure 2mm to 8mm in length however the leg span is much larger and can measure 10mm to 170mm in length.
They are better known as “harvestmen” or “daddy longlegs” These Arachnids that are harmless to people and are known for their exceptionally long walking legs, compared to their body size. There are over 6,500 species of Opiliones world wide which are divided into four suborders: Cyphophthalmi, Eupnoi, Dyspnoi and Laniatores.
Well preserved fossils have been found in the Rhynie cherts of Scotland which are around 410 million years old. Those fossils look exceptionally modern, suggesting that the basic structure of the “crab harvestmen” has not changed much since then.

In more advanced species, the first five abdominal segments are often fused into a dorsal shield called the scutum, which is normally fused with the carapace. At times this shield is only present in the male spiders. The two hindmost abdominal segments may be reduced or separated in the middle on the surface to form two plates lying next to each other. The second pair of legs is longer than the others and works as antennae.
They have a single pair of eyes in the middle of their heads, oriented sideways. They have a pair of prosomatic scent glands that secrete a foul smelling fluid when disturbed. Harvestmen do not have spinnerets and do not possess poison glands.
They breathe through tracheae. Between the base of the fourth pair of legs and the abdomen is a pair of spiracles, one opening on each side.
In more active species, spiracles are also a pair of spiracles found on the tibia of the legs.
Opiliones have a gonopore on the ventral cephalothorax, and copulation is direct, as the male has a penis while the female has an ovipositor.
Most species are short lived living around 1 year or less. Most species are omnivorous, eating primarily small insects and all kinds of plant material and fungi while some are scavengers of the decays of any dead animal or fecal material particularly bird fecal.
Most are nocturnal and fawn to brown, although many can be diurnal which have vivid patterns in yellow, green and black with varied reddish and blackish mottling and reticulated markings.
Pholcidae or Daddy Long Legs also known as cellar spiders, have long filiform like legs just like Opiliniones but the body is divided in two sections. They build irregular webs in and around buildings and can be found beneath furniture. The spider was accidently introduced to Australia from Europe and is now one of the most common spiders in suburbia. The latest revision of the genus has increased the number of species from tow to 16.

Salticidae
The jumping spiders as their common name suggests are excellant jumpers. They do not build webs and usually have excellent camouflage blending into the foliage or bark of trees and shrubs they frequent. The 3mm to 12mm in length spiders have short strong legs and seem to have personalities bestowed upon them.
Their large eyes have excellent eye sight and assist in the location of food hiding amongst the foliage. Most are daylight hunters, using their excellent vision to detect and calculate distance, before suddenly leaping onto their prey.
Most of the tropical species are brilliantly coloured in bright green bases with yellow orange and red markings which probably assists in luring pollen insect closer enough to be pounced upon.
The males lead the way in colour, brilliance and hair patterns. They use these adornments to impress the females during their elaborate courtship displays. Members of the Maratus genus are known as the peacocks of the spider world.
It is believed that the male Maratus volans clad in red, blue and black has the most spectacular courtship display of all spiders and contrasts vividly to his prospective partner dressed in fawn and brown. He expands and raises the lateral flaps on his abdomen so that they form a white circular aurora around the field of blazing colour which rises up above his brilliantly coloured carapace. He then parades around her keeping his upper abdomen facing towards her head at all times.
The tropic scene is also the home of the most adapted hunter of all Portia fimbriata. Covered in lichen like hair tufts and small spines, this jumper can stalk prey, ambush prey, is adept at invading the webs of other spiders and imitates the strategies of other spiders to it preys on.
Jumping spiders are diurnal and on warm days can be found on most types of vegetation.
Theridiidae or combfooted spiders also known as house spiders and have a glabrous globular abdomen. These spiders build loose, irregular webs. They have shiny, slender legs, with a small cephalothorax and a larger orbicular to ovoidal-orbicular abdomen; the colour can range from a brown or reddish plum to satiny black. The abdomen often has white or beige spots, a frontal crescent, and sometimes, small red spots or a thin red line. Most measure 8mm to 20 mm in body length.
The spiders build a tangled looking web with sticky lines (a gum footed web) under rocks, timber, and bark or near cave entrances. Their common name refers to them being found in any dark, sheltered, place around the house or garden especially among old furniture and junk stored in sheds and garages, under garden benches, around compost bins, in upturned flower pots and the like.
Thomisidae or tarantulas, crab spiders or bark spiders have the first pair, second pair of legs or two pairs longer than the back pairs and like crabs often walk sideways or backwards. They do not build webs.
There are around 2166 described species making it one of the most common spider families in the world.
Most Thomisidae are found on vegetation or in leaf litter or on the forest floor while others have a close association with flowers. Their compact, globose bodies have relatively short, stocky legs and a crab like, lateral posture where the legs extend sidewise and the femora are twisted so that the front surface faces upwards. This makes them generally easy to recognize.
The first pair of legs is often armed with heavy raptorial spines that are used extensively in securing the prey once it is captured. Thomisidae have eight eyes of which the lateral eyes are typically situated on prominent pale tubercles and are often larger than the median eyes. Thomisidae in the genus Coriarachne can flatten their bodies and live in crevices beneath loose tree bark but can also be found on fence posts and wooden buildings. The flower inhabiting Thomisidae in the genre Misumena, Minumenoides & Misumenops are all ambush pollinating insects. Some of these spiders have the ability to slowly change their color to blend into the background.
The red back spider has a reputation of being extremely dangerous when in fact, while dangerous is usually non fatal.
- Intense localised pain, with swelling and sweating starting five minutes after the bite
- Pain and swelling may be experienced over the body after about half an hour
- Headache, nausea and vomiting may occur after one hour – profuse sweating is common.
The following first aid is recommended:
- Keep patient calm
- Apply ice packs (such as a packet of frozen peas wrapped in a tea towel) to minimise pain and swelling
- Seek medical aid – anti-venene will be administered if symptoms are severe
- Do not use a tourniquet or restrictive bandage, or excise or incise the bite. https://museum.wa.gov.au/

Non spider Arachnids
Palpigradi or Palpigrads or “microwhip scorpions” are small species which are smaller than 3mm in length and are found world wide except for the Polar Regions. The thin, pale, segmented carapace terminates in a 15 segmented, whip like flagellum. The flagellum is kept vertical when moving.
The carapace is divided into two plates between the third and fourth leg set. They have no eyes. Some species have three pairs of book lungs, while others have no respiratory organs at all.
They are predatory feeding on minute insects in their ecosystems. Their sexual habits and courtships are unknown. The females lay only a few relatively large eggs. They live in moist environments under partially buried stones and rocks.

Western Australian Museum
Thelyphonida or whip scorpions are small species which measure 3mm to 85mm in length and are found world wide in tropical and subtropical areas of the world. They usually live in moist underground burrows that they dig with their pedipalps. They may also burrow under logs, rotting wood and rocks avoiding the sun light.
They have six legs for walking while the front pair is modified into antennae like sensory organs. Many species also have very large scorpion like pedipalps. They have one pair of eyes at the front of the cephalothorax and three on each side of the head. Thelyphonida do not have poison glands or a stinger on the abdomen. Most possess special glands near the rear of their abdomen that can spray a combination of acetic acid and octanoic acid when they are aggravated or threatened. Some species spray formic acid while others spray a chlorine compound. The segmented carapace terminates in a long many segmented, semi rigid flagellum.

http://commons.wikimedia.org/wiki/File: Whip scorpion.
Whip scorpions are nocturnal, carnivores, feeding mostly on insects especially roaches and crickets but at times on worms and slugs. The prey is crushed between special teeth on the inside of the trochanters which are found on the second segment of the front legs.
Males secrete a sperm sac, which is transferred to the female. Up to 35 eggs are laid in a burrow, within a mucous membrane that preserves moisture. The females remain with the eggs and do not eat. The translucent to opaque, white young hatch from the eggs, climb onto their mother’s back and attach themselves there with special suckers. After the first molt, they look like miniature whip scorpions, and leave the burrow; the mother dies soon after. Thelyphonida go through three molts in about three years.
Schizomida refers to the “split or cleaved middle” which refers to the cephalothorax being divided into two separate plates. Schizomids use three pairs of legs for walking while the front pair of legs is modified as sensory organs. They also have large well developed pedipalps just behind the sensory legs. They differ from Thelyphonida in that it has a short stout 4 rigid, segmented appendage.
Schizomida prefer to live in the top layer of soil and beneath thick forest litter. Schizomids present the prosoma covered by a large protopeltidium and smaller, paired, mesopeltidia and metapeltidia. They do not possess eyes. The opisthosoma is a smooth oval of 12 recognizable somites. The first is reduced and forms the pedicel. The last three are much constricted, forming the pygidium.

Amblypygids are also known as tailless whip scorpions or cave spiders. To date 5 families, 17 genera and 136 species have been described. Fossilised amblypygids have been found dating back to the Carboniferous period. They are found in tropical and subtropical regions worldwide. Some species are subterranean while the remainder are nocturnal. They usually live under logs, bark, stones, or leaves preferring ecosystems which are moist or humid. Amblypygids may range from 5mm to 40 mm. Their bodies are broad and flattened with the first pair of legs are modified sensory organs. (Compare solifugids, uropygids, and schizomids.) These very thin modified legs can extend several times the length of body. They have no silk glands or venomous fangs, but often have very prominent pincer like pedipalps. Amblypygids often move about sideways on their six walking legs, with one “whip” pointed in the direction of travel while the other probes are on either side of them. Prey are located with these “flagella like whips”, captured with pedipalps and then torn to pieces with the chelicerae.

Amblypygids, particularly the species Phrynus marginemaculatus and Damon diadema, are thought to be one of the few species of arachnids that show signs of social behavior. Research conducted at Cornell University by entomologists suggests that mother amblypygids comfort their young by gently caressing the offspring with her feelers. Further, when two or more siblings were placed in an unfamiliar environment, such as a cage, they would seek each other out and gather back in a group.
Scorpiones
The true scorpions have bodies that are divided into two sections, cephalothorax and abdomen. The Pedipalps are very large and end with large chelae. They are characterized by a curved metasoma (tail) comprising of six segments. The first segment contains the sexual organs as well as a pair of vestigial and modified appendages forming a structure called the genital operculum.
The second segment bears a pair of feather like sensory organs known as the pectines. The pectines are still controversial but according to experiments conducted by Kleber Del-Claro of the University of Federal de Uberlandia there is a strong correlation to hunting techniques and success in hunting.
The final four segments each contain a pair of book lungs. The mesosoma is armored with chitinous plates, known as tergites on the upper surface and sternites on the lower surface.
The final segment contains the scorpion’s anus and bears the telson (the sting). The telson, in turn, consists of the vesicle, which holds a pair of venom glands and the hypodermic aculeus, the venom injecting barb. The abdomen’s front half, the mesosoma, is made up of six segments.
The cuticle of scorpions is covered with hairs in some places that act like balance organs. An outer layer that makes them fluorescent green under ultraviolet light is called the hyaline layer. Newly molted scorpions do not glow until after their cuticle has hardened. The fluorescent hyaline layer can be intact in fossil rocks that are hundreds of millions of years old.
Scorpiones are fossorial, nocturnal arachnids finding shelter during the day in the relative cool locations like in underground holes or the underside of rocks and logs and coming out at night to hunt and feed. Scorpions prefer to live in areas where the temperatures range from 20 to 37 °C, but may survive in the temperature range of 14–45 °C.
Scorpions are opportunistic predators of small arthropods and insects. They use their chela (pincers) to catch the prey initially. Depending on the toxicity of their venom and size of their claws, they will then either crush the prey or inject it with neurotoxic venom. The neurotoxins consist of a variety of small proteins as well as sodium and potassium cations, which serve to interfere with neurotransmission in the victim. Scorpions use their venom to kill or paralyze their prey so that it can be eaten. It is fast acting, allowing for effective prey capture. Scorpion venoms are optimized for action on other arthropods and therefore most scorpions are relatively harmless to humans.
The stings from the Australian species produce only local effects (such as pain, numbness or swelling). Interestingly though the scorpion that is responsible for the most human deaths is the Androctonus australis, or fat-tailed scorpion of North Africa. (australis is Greek for the original Great South Land that refers to Africa not Australia.) The toxicity of A. australis’s venom is roughly half that of the death stalker (Leiurus quinquestriatus), but since A. australis injects quite a deal more venom it is the most deadly to humans. Scorpions have two types of venom. Translucent, weaker venom designed to stun only, and opaque, more potent venom designed to kill heavier threats.
Scorpions are viviparous bearing live young which are carried about on the females back until the young have undergone at least one moult. They undergo between five and seven moults to reach maturity. Scorpions have quite variable life spans. The age range appears to be between 4 to 25 years.
Fossil records of scorpions date from the Carboniferous Period and in marine Silurian deposits some 450 to 425 million years ago. It is believed that they have an oceanic origin, with gills and a claw like appendage that enabled them to hold onto rocky shores or seaweed.

Pseudoscorpions are small arthropods with a flat, pear-shaped body and pincers that resemble those of scorpions. They range from 2mm to 8mm in length. The opisthosoma is made up of twelve segments, each guarded by plate like tergites above and sternites below. The abdomen is short and obtuse at the rear, rather than extending into a segmented tail and stinger like true scorpions. The colour of the body can be yellowish-tan to deep-brown, with the paired claws often a contrasting colour. They may have two, four or no eyes. They have two very long pedipalps with palpal chelae that strongly resemble the pincers found on a scorpion.
The pedipalps generally consist of an immobile “hand” and “finger” with a separate movable finger controlled by an adductor muscle. There is a venom gland and duct, usually located in the mobile finger; the poison is used to capture and immobilize the pseudo scorpion’s prey.
There are several characteristics that resemble their close relatives the true spiders. During digestion, they excrete a mildly corrosive fluid over the prey, liquefying remains prior to ingesting it. Pseudoscorpions also spin silk from a gland in their jaws to make disk shaped cocoons for mating, molting, or waiting out cold weather. Another trait they share with spiders is that they breathe through spiracles. Most spiders have one pair of spiracles, and one of book lungs; however pseudoscorpions do not possess book lungs.
Pseudoscorpions prey on clothes moth larvae, carpet beetle larvae, booklice, ants, mites, and small flies. They may sometimes be found feeding on mites under the wing covers of certain beetles.
There are more than 2,000 species of pseudoscorpions recorded. They range worldwide with most being found in the tropics and subtropics but they also extend to near the Polar Regions. The fossil record of pseudoscorpions dates back over 380 million years, to the Devonian period.
Pseudoscorpions have elaborate mating dances with the males of some pseudoscorpion species enticing a female over a spermatophore he ahs previously laid upon the surface. In other species, the male also pushes the sperm into the female genitals using his forelegs. The female carries the fertilized eggs in a brood pouch attached to her abdomen, and the young ride on the mother for a short time after they hatch. Up to two dozen young are hatched in a single brood; there may be more than one brood per year. The young go through three molts over the course of several years before reaching adulthood. Adult pseudoscorpions live 2 to 3 years. They are active in the warm months of the year, overwintering in silken cocoons when the weather is too cold.

Ricinulei
The hooded tick spiders measure 5mm to10 mm long. Their most notable feature is a “sliding hood” that can be raised and lowered over the head and mouth parts and the chelicerae. Ricinulei have no eyes. The pedipalps end in pincers that are small relative to their bodies, when compared to those of the related orders of scorpions and pseudoscorpions. The heavy bodied abdomen tapers to a narrow pedicel where it attaches to the prosoma. The trachea directly exchanges gasses to the body.
There are presently 57 species of Ricinuleids described worldwide mainly from the tropical and subtropical regions. They are all in a single family that contains 3 genera.
Ricinulei
These are predators, feeding on other small arthropods.
In males, the third pair of legs is modified to form copulatory organs. Malpighian tubules and a pair of coxal glands make up the excretory system. Beyond this little is known about their sexual habits though the males have been observed using their modified third leg to transfer a spermatophore to the female. The single egg; of those species observed, is carried under the mother’s hood, until the young hatch into six-legged “larva”. They later moult into their adult forms. Ricinulei live in dark moist locations under forest litter.
Solifugae or solifugids,
The wind or whip scorpions resemble pseudoscorpions but are bigger growing from 8mm to 20mm in length. The pedipalps end with an adhesive organ. The most distinctive feature of Solifugae is their large chelicerae. Each of the two chelicerae comprise of two articles which form powerful pincers. Each article bears a variable number of teeth. In the species of the order Amblypygi they are at least 0.5mm length while the chelicerae instead have small pedipalps which are strong, thorny and without an adhesive organ. The fore legs are thin and extremely long.
There are about 900 species of Solifugae which is derived from Latin, and means those that flee from the sun. Most Solifugae live in dry monsoonal, tropical or semitropical regions where they inhabit warm dry or semi arid and arid habitats with some species living in grassland or open woodland habitats.
The males in all families except Eremobatidae possess a flagellum on the basal article of the chelicera. Solifugae also have long pedipalps, which function as sense organs similar to insects’ antennae and give the appearance of the two extra legs. Pedipalps terminate in reversible adhesive organs.
These are carnivorous or omnivorous, with most species feeding on termites, beetles, and other small arthropods, however, they have been recorded eating larger prey, such as skinks. Prey is located with the pedipalps and killed and cut into pieces by the chelicerae. The prey is then liquefied and the liquid ingested through the pharynx.
Reproduction can involve direct or indirect sperm transfer. When it is indirect the male emits a spermatophore on the ground and then inserts it with his chelicerae in the female’s genital pore.

Crustaceans
I have included a small section here on Crustaceans because many individual species live in the tidal zones between land and water with some species living most their lives above the high water make and they are the ancestors to all the living terrestrial arthropods.
Characteristics
The name “crustacean” dates from the earliest works to describe animals, including those of Pierre Belon and Guillaume Rondelet, but the name was not used by some later authors, including Carl Linnaeus, who included crustaceans among the “Aptera” in his Systema Naturae. The earliest nomenclaturally valid work to use the name “Crustacea” was Morten Thrane Brünnich’s Zoologiæ Fundamenta in 1772, although he also included chelicerates in the group.
The subphylum Crustacea comprises almost 67,000 described species, which is thought maybe only 1mm to 10mm of the total number as the majority of species remain as yet undiscovered or unnamed. Although most crustaceans are small, their morphology varies greatly and includes both the largest arthropod in the world which is from Japan. The Japanese spider crab Macrocheira kaempferi has a leg span of 3800mm while the smallest Stygotantulus stocki measures just 1micrometre in length. Despite their diversity of size and form, crustaceans are united by the special larval form known as the nauplius.
The exact relationships of the Crustacea to other taxa are not completely settled and studies based on morphology led to the Pancrustacea hypothesis, in which Crustacea and Hexapoda in which insects and their allies are the terrestrial sister groups. More recent studies using DNA sequences suggest that Crustacea is paraphyletic, with the hexapods nested within a larger Pancrustacea clade. Although the classification of crustaceans has been quite variable research is on classification is still on going. The system presently used is by Martin and Davis for the major part largely supersedes earlier works. Mystacocarida and Branchiura are treated as part of the Maxillopoda and are at times treated as their own classes. Six classes are usually recognized.
The Body structures of crustacean are composed of 3 body segments which are known as the cephalon or head, the thorax, and the pleon or abdomen.
The head and thorax may be fused together to form a cephalothorax, which may be covered by a single large carapace. The crustacean’s body is protected by the hard exoskeleton, which must be moulted periodically for the animal to grow. The shell around each somite can be divided into a dorsal tergum, ventral sternum and a lateral pleuron. Various parts of the exoskeleton may be fused together.
Each somite, or body segment can bear a pair of appendages on the segments of the head, these include two pairs of antennae, the mandibles and maxillae. The thoracic segments bear the legs. The legs may be specialized as pereiopods for walking or maxillipeds for feeding. The abdomen bears pleopods, and ends in a telson, which bears the anus, and is often flanked by uropods to form a thin leathery type tail. The number and variety of appendages varies greatly.
Crustacean appendages are typically biramous, meaning they are divided into two parts; this includes the second pair of antennae, but not the first, which is usually uniramous, the exception being in the Class Malacostraca where the antennules may be generally biramous or even triramous. It is unclear whether the biramous condition is a derived state which evolved in crustaceans, or whether the second branch of the limb has been lost in all other groups. Trilobites, for instance, also possessed biramous appendages.
The main body cavity is an open circulatory system, where blood is pumped into the haemocoel by a heart located near the dorsum. Malacostraca have haemocyanin as the oxygen carrying pigment, while Copepods, Ostracods, Barnacles and Branchiopods have haemoglobins. The alimentary canal consists of a straight tube that often has a gizzard like “gastric mill” for grinding food and a pair of digestive glands that absorb food. This structure goes in a spiral format. Structures that resemble and function like ancient kidneys are located near the antennae. A brain exists in the form of ganglia close to the antennae, and a collection of major ganglia is also found below the gut.
Crustaceans consist of 6 major Classes and 52 orders.
<>
Class Members Orders Photo
Branchiopoda
brine shrimp
Cladocera
Triops
Anostraca
Lipostraca
Notostraca
Laevicaudata
Spinicaudata
Cyclestherida
Cladocera
Daphnia pulex (Cladocera)
Remipedia
Nectiopoda
Speleonectes tanumekes (Speleonectidae)
Cephalocarida
horseshoe shrimp
Brachypoda
Maxillopoda
barnacles
copepods
Calanoida
Pedunculata
Sessilia
c. 20 others
Chthamalus stellatus (Sessilia)
Ostracoda
ostracods
Myodocopida
Halocyprida
Platycopida
Podocopida
Cylindroleberididae
Malacostraca
crabs
lobsters
shrimp
krill
mantis shrimp
woodlice
sandhoppers
etc. Decapoda
Isopoda
Amphipoda
Stomatopoda
c. 12 others
Gammarus roeseli (Amphipoda
The Ecology of the majority of crustaceans are aquatic, living in either marine or freshwater environments with a few groups adapting to life on land, such as terrestrial crabs, terrestrial hermit crabs, and woodlice. Marine crustaceans are as varied in the oceans as insects are on land. Most crustaceans are free moving although a few species are parasitic and live attached to their hosts. These include sea lice, fish lice, whale lice, tongue worms, and Cymothoa exigua, all of which may be referred to as “crustacean lice”. Adult barnacles live a sessile life attached by the head to the substrate. Some Branchiurans are able to withstand rapid changes of salinity and are able to switch hosts from marine to non marine species. Krill are the bottom layer and the most important part of the food chain in the southern waters of the Antarctica where they are fed upon by most animal communities including the giant whales. Some crustaceans are very invasive such as the Chinese mitten crab, Eriocheir sinensis, and the Asian shore crab, Hemigrapsus sanguineus both of which would be extremely damaging to the Australia ecosystems if they managed to find there way into Australian coastal waters.
Life Cycles
The majority of crustacean have separate sexes, and reproduce sexually. A small number are hermaphrodites which include the Barnacles, remipedes and Cephalocarida and it is thought that a few may even change sex during the course of their lives. Parthenogenesis is also widespread among crustaceans, where viable eggs are produced by a female without needing fertilization by a male. This occurs in many branchiopods, some ostracods, some isopods, and certain “higher” crustaceans, such as the Marmorkrebs crayfish.
The egg distribution and protection is as varied as the Crustaceans themselves. In many groups of crustaceans, the fertilized eggs are simply released into the water or water current, while others have developed a number of mechanisms for holding on to the eggs until they are ready to hatch. Peracarids, Notostracans, Anostracans, and many Isopods form a brood pouch from the carapace and thoracic limbs. Female Branchiura do not carry eggs in external ovisacs but attach them in rows to rocks and other objects. Most Leptostracans and krill carry the eggs between their thoracic limbs while some Copepods carry their eggs in special thin walled sacs, while others have them attached together in long, tangled strings.
In many male decapods, the first and at times the second pair of pleopods are specialized to transfer sperm to the females. Many terrestrial crustaceans like the Christmas Island red crab mate seasonally and return to the sea to release the eggs. Others, such as woodlice, lay their eggs on land, albeit in wet conditions. Most females decapods retain the eggs attached to the pleopods and are wrapped between the pleopod and the under body until they hatch into free swimming larvae.
Crustaceans exhibit a number of larval forms, of which the earliest and most characteristic is the nauplius. The nauplii have three pairs of appendages, all emerging from the head, and a single naupliar eye. In most groups, there are further larval stages known as the zoea. It follows the nauplius stage and precedes the post larva. Zoea larvae swim with their thoracic appendages, as opposed to nauplii, which use cephalic appendages, and megalopa, which use abdominal appendages for swimming. It often has spikes on its carapace, which are thought at this stage to assist these small organisms in maintaining directional swimming.
In many decapods, the zoea is the first larval stage. In some cases, the zoea stage is followed by the mysis stage, and in others, by the megalopa stage, depending on the crustacean group involved.
Insects
Characteristics Before we start insects remember insects are more diverse than spiders. The following description was adapted from Wikipedia http//www.en.wikipedia.org/insects
The Body structures of insects have been extensively studied with segmented bodies supported by exoskeletons. The hard outer covering is made mostly of chitin the same material in which finger nails are made. The segments of the body are organized into three distinctive but interconnected segments, or tagmata, the head, the thorax and the abdomen.
The head supports a pair of sensory antennae, a pair of compound eyes, and, if present, one to three simple eyes (or ocelli) (2 & 3) and three sets of variously modified appendages that form the mouthparts. The thorax has six segmented legs with one pair each on the prothorax, mesothorax and the metathorax. The thorax supports none, one pair of wings or two pairs of wings. The abdomen consists of 11 segments, though in a few species of insects, these segments may be fused together or reduced in size. The abdomen also contains most of the digestive, respiratory, excretory and reproductive internal structures. Considerable variation and many adaptations in the body parts of insects occur, especially wings, legs, antenna and mouthparts which are vital in the identification of the various classes, orders, families, genre and species.
The head is enclosed in a hard, heavily sclerotized, unsegmented, exoskeletal head capsule, or epicranium, which contains most of the sensing organs, including the antennae, compound and simple eyes, and mouthparts. Of all the insect orders, Orthoptera displays the most features found in other insects, including the sutures and sclerites. Here, the vertex, or the apex (dorsal region), is situated between the compound eyes for insects with a hypognathous and opisthognathous head. In prognathous insects, the vertex is not found between the compound eyes, but rather, where the ocelli are normally. This is because the primary axis of the head is rotated 90° to become parallel to the primary axis of the body. In some species, this region is modified and assumes a different name.
The thorax is a tagma composed of three sections, the prothorax, mesothorax and the metathorax. The anterior segment, closest to the head, is the prothorax, with the major features being the first pair of legs and the pronotum. The middle segment is the mesothorax, with the major features being the second pair of legs and the anterior wings. The rear segment, abutting the abdomen, is the metathorax, which features the third pair of legs and the posterior wings. Each segment is dilineated by an intersegmental suture. Each segment has four basic regions. The dorsal surface is called the tergum (or notum) to distinguish it from the abdominal terga. The two lateral regions are called the pleura (singular is pleuron) and the ventral aspect is called the sternum. In turn, the notum of the prothorax is called the pronotum, the notum for the mesothorax is called the mesonotum and the notum for the metathorax is called the metanotum. Continuing with this logic, the mesopleura and metapleura, as well as the mesosternum and metasternum, are also used.
The abdomen: is the largest tagma of the insect, which typically consists of 11 to 12 segments and is less strongly sclerotized than the head or thorax. Each segment of the abdomen is represented by a sclerotized tergum and sternum. Terga are separated from each other and from the adjacent sterna or pleura by membranes. Spiracles are located in the pleural area. Variation of this includes the fusion of terga or terga and sterna to form continuous dorsal or ventral shields or a conical shaped tube. Some insects bear a sclerite in the pleural area called a laterotergite. Ventral sclerites are sometimes called laterosternites. During the embryonic stage of many insects and the post embryonic stage of primitive insects there are 11 abdominal segments. In modern insects there is a tendency toward reduction in the number of the abdominal segments, but the primitive number of 11 is maintained during embryogenesis. Variation in abdominal segment number is considerable. If the Apterygota are considered to be indicative for pterygotes then confusion is seen as adult Protura have 12 segments while the Collembola have 6. The orthopteran family Acrididae has 11 segments, and a fossil specimen of Zoraptera has a 10 segmented abdomen.
The Exoskeleton of an insect’s outer skeleton or cuticle is made up of two layers: the epicuticle, which is a thin and waxy water resistant outer layer and contains no chitin, and a lower layer called the procuticle. The procuticle is chitinous and much thicker than the epicuticle and has two layers: an outer layer known as the exocuticle and an inner layer known as the endocuticle. The tough and flexible endocuticle is built from numerous layers of fibrous chitin and proteins, criss crossing each other in a sandwich pattern, while the exocuticle is rigid and hardened. The exocuticle is greatly reduced in many soft bodied insects especially during their larval stages.
Insects are the only invertebrates that have evolved to the stage of flying, and this has played an important role in their success as mobility enhances their ability to travel long distance and to populate new localities such as islands. Their muscles are able to contract multiple times for each single nerve impulse, allowing the wings to beat faster than would ordinarily be possible. Having their muscles attached to their exoskeletons is more efficient and allows more muscle connections. Crustaceans also use the same method, though spiders use hydraulic pressure to extend their legs, a system inherited from their pre-arthropod ancestors. Unlike insects, though, most aquatic crustaceans are biomineralized with calcium carbonate extracted from the water.
The nervous system of an insect can be divided into a brain and a ventral nerve cord. The head capsule is made up of six fused segments, each with either a pair of ganglia, or a cluster of nerve cells outside of the brain. The first three pairs of ganglia are fused into the brain (5), while the three following pairs are fused into a structure of three pairs of ganglia under the insect’s esophagus, called the subesophageal ganglion.
The thoracic segments have one ganglion on each side, which are connected into a pair, one pair per segment (19). This arrangement is also seen in the abdomen but only in the first eight segments. Many species of insects have reduced numbers of ganglia due to fusion or reduction. Some cockroaches have just six ganglia in the abdomen, whereas the wasp Vespa crabro has only two in the thorax and three in the abdomen. Some insects, like the house fly Musca domestica, have all the body ganglia fused into a single large thoracic ganglion.
At least a few insects have nociceptors, cells that detect and transmit sensations of pain. This was discovered in 2003 by studying the variation in reactions of larvae of the common fruit fly Drosophila melanogaster. to the touch of a heated probe and an unheated one. The larvae reacted to the touch of the heated probe with a stereotypical rolling behavior that was not exhibited when the larvae were touched by the unheated probe. Although nociception has been demonstrated in insects, there is no consensus that insects feel pain consciously.
The digestive System of an insect uses its digestive system to extract nutrients and other substances from the food it consumes.[40] Most of this food is ingested in the form of macromolecules and other complex substances like proteins, polysaccharides, fats and nucleic acids. These macromolecules must be broken down by catabolic reactions into smaller molecules like amino acids and simple sugars before being used by cells of the body for energy, growth, or reproduction. This break-down process is known as digestion.
The main structure of an insect’s digestive system is a long enclosed tube called the alimentary canal, which runs lengthwise through the body. The alimentary canal directs food unidirectional from the mouth to the anus (17a). It has three sections, each of which performs a different process of digestion. In addition to the alimentary canal, insects also have paired salivary glands and salivary reservoirs. These structures usually reside in the thorax, adjacent to the foregut.
The salivary glands (30) in an insect’s mouth produce saliva. The salivary ducts lead from the glands to the reservoirs and then forward through the head to an opening called the salivarium, located behind the hypopharynx. By moving its mouthparts the insect can mix its food with saliva. The mixture of saliva and food then travels through the salivary tubes into the mouth, where it begins to break down. Some insects, like flies, have extra oral digestion. Insects using extra oral digestion expel digestive enzymes onto their food to break it down. This strategy allows insects to extract a significant proportion of the available nutrients from the food source. The gut is where almost all of insects’ digestion takes place. It can be divided into the foregut, midgut and hindgut or intestinal tract .
Digestion starts in buccal cavity (mouth) as partially chewed food is broken down by saliva from the salivary glands. As the salivary glands produce fluid and carbohydrate digesting enzymes; mostly amylases, the pharynx muscles pump fluid into the buccal cavity, lubricating the food like the salivarium does, and helping blood feeders, and xylem and phloem feeders.
The first section of the alimentary canal is the foregut, or stomodaeum. The foregut is lined with a cuticular lining made of chitin and proteins as protection from tough food. The foregut includes the buccal cavity (mouth), pharynx, esophagus and crop and proventriculus (any part may be highly modified) which both store food and signify when to continue passing onward to the midgut.
From the pharynx food moves to the esophagus, which could be just a simple tube passing it on to the crop and proventriculus, and then onward to the mid gut, as in most insects. Alternately, the foregut may expand into a very large crop and proventriculus, or the crop could just be a diverticulum, or fluid filled structure, as in some Diptera species.
Once food leaves the crop, it passes to the midgut (13), also known as the mesenteron, where the majority of digestion takes place. Microscopic projections from the mid gut wall, called microvilli, increase the surface area of the wall and allow more nutrients to be absorbed; they tend to be close to the origin of the mid gut. In some insects, the role of the microvillus and where it is located may vary. For example, specialized microvillus producing digestive enzymes may more likely be near the end of the mid gut, and absorption near the origin or beginning of the mid gut.
In the hindgut (16 & 17 intestine or late hindgut), or proctodaeum, undigested food particles are joined by uric acid to form fecal pellets. The rectum absorbs 90mm of the water in these fecal pellets, and the dry pellet is then eliminated through the anus, completing the process of digestion. The uric acid is formed using hemolymph waste products diffused from the Malpighian tubules (20). The uric acid is then emptied directly into the alimentary canal, at the junction between the mid gut and hindgut. The number of Malpighian tubules possessed by a given insect varies between species, ranging from only two tubules in some insects to over 100 tubules in others.
The reproductive system of female insects, consist of a pair of ovaries, accessory glands, one or more spermathecae (15a), and ducts connecting these parts. The ovaries are made up of a number of egg tubes, called ovarioles, which vary in size and number by species. The number of eggs that an insect is able to make, varies by the number of ovarioles with the rate that eggs can be develop being also influenced by ovariole design.
Female insects are able make eggs, receive and store sperm, manipulate sperm from different males, and lay eggs. Accessory glands or glandular parts of the oviducts produce a variety of substances for sperm maintenance, transport and fertilization, as well as for protection of eggs. They can produce glue and protective substances for coating eggs or tough coverings for a batch of eggs called oothecae. Spermathecae are tubes or sacs in which sperm can be stored between the time of mating and the time an egg is fertilized.
For males, the reproductive system is the testis, suspended in the body cavity by tracheae and the fat body. Most male insects have a pair of testes, inside of which are sperm tubes or follicles that are enclosed within a membranous sac. The follicles connect to the vas deferens by the vas efferens, and the two tubular vas deferentia connect to a median ejaculatory duct that leads to the outside. A portion of the vas deferens is often enlarged to form the seminal vesicle, which stores the sperm before they are discharged into the female. The seminal vesicles have glandular linings that secrete nutrients for nourishment and maintenance of the sperm. The ejaculatory duct is derived from an invagination of the epidermal cells during development and, as a result, has a cuticular lining. The terminal portion of the ejaculatory duct may be sclerotized to form the intromittent organ known as the aedeagus. The remainder of the male reproductive system is derived from embryonic mesoderm, except for the germ cells, or spermatogonia, which descend from the primordial pole cells very early during embryogenesis.
The tube like heart of the mosquito Anopheles gambiae extends horizontally across the body, interlinked with the diamond shaped wing muscles and surrounded by pericardial cells.
Insect respiration is accomplished without lungs. Instead, the insect respiratory system uses a system of internal tubes and sacs through which gases either diffuse or are actively pumped, delivering oxygen directly to tissues that need it via their trachea (8). Since oxygen is delivered directly, the circulatory system is not used to carry oxygen, and is therefore greatly reduced. The insect circulatory system has no veins or arteries, and instead consists of little more than a single, perforated dorsal tube which pulses peristaltically. Toward the thorax, the dorsal tube (14) divides into chambers and acts like the insect’s heart. The opposite end of the dorsal tube is like the aorta (8) of the insect circulating the hemolymph, arthropods’ fluid analog of blood, inside the body cavity. Air is taken in through openings on the sides of the abdomen called spiracles.
The respiratory system is an important reason that limits the size of insects. As insects get bigger, this type of oxygen transport gets less efficient and thus the heaviest insect currently weighs less than 100 grams. When atmospheric oxygen levels increased during the late Paleozoic, larger insects evolved like the giant dragonflies with wingspans of around 600mm.
There are many different patterns of gas exchange demonstrated by different groups of insects. Gas exchange patterns in insects can range from continuous and diffusive ventilation, to discontinuous gas exchange. During continuous gas exchange, oxygen is taken in and carbon dioxide is released in a continuous cycle. In discontinuous gas exchange, however, the insect takes in oxygen while it is active and small amounts of carbon dioxide are released when the insect is at rest. Diffusive ventilation is simply a form of continuous gas exchange that occurs by diffusion rather than physically taking in the oxygen. Some species of insect that are submerged also have adaptations to aid in respiration. As larvae, many insects have gills that can extract oxygen dissolved in water, while others need to rise to the water surface to replenish air supplies which may be held or trapped in hair like structures along the legs or around the anus.
Insect ecology varies greatly on whether the insect is solitary or socially bound and how it interacts with its surrounding environment or ecosystem. Insects play one of the most important roles in their ecosystems, which include soil aeration, dung burial and consumption, pest control, pollination and wildlife nutrition.
Insects are mostly soft bodied, fragile and almost defenseless compared to other forms of life. The immature stages are more vulnerable than the adults; at every stage of their short lives, being exposed to carnivores and parasites. For this reason insects then have evolved a wide variety of defense strategies to avoid becoming a meal. These include camouflage, mimicry, toxicity and active defense.
Camouflage is the most prominent defense strategy diploid by insects. It involves the use of coloration and or shape to blend into its surrounding environment. This sort of protective coloration is common and widespread among beetle families, especially those that feed on wood or vegetation, such as many of the leaf beetles or weevils. Some species scales are roughly and irregularly constructed with the use of various coloured scales or hairs to cause the beetles to resemble bird dung or other inedible objects. Many of those that live on the large surface environments blend in with the coloration of the soil, rocks, trunk or bark. Most stick insects (Phasmid specie) are known for effectively replicating the forms of sticks and leaves, and the bodies of some species like Palophus centaurus are covered in moss or lichen outgrowths that supplement their disguise. Some species have the ability to change color as their surroundings change Bostra scabrinota.
The Phasmatodea have developed several further behavioral adaptations to aid their camouflage. Some insects along with the Phasmatodea use crypsis, where they perform a rocking motion swaying the body from side to side. This is considered to be a reflection of the movement of leaves or twigs swaying in the breeze.
Another method by which most stick insects deploy is catalepsy to avoid predation and resemble twigs is by feigning death, where the insect enters a motionless state that can be maintained for a long period. The nocturnal feeding habits of adults also aids Phasmatodea in remaining concealed from predators.
Mimicry is another defense that often uses color or shape to deceive potential enemies. A number of longhorn beetles in the family Cerambycidae; bear a striking resemblance to wasps, which helps them avoid predation even though the beetles are in fact harmless.
Batesian and Müllerian mimicry complexes are commonly found in Lepidoptera or butterflies. Genetic polymorphism and natural selection give rise to otherwise edible species gaining a survival advantage by resembling inedible species. Such a mimicry complex is referred to as Batesian and is most commonly known by the mimicry by the Limenitidine Viceroy butterfly of the inedible Danaine Monarch butterfly. Later research has discovered that the Viceroy is, in fact more toxic than the Monarch and this resemblance should be considered as a case of Müllerian mimicry.
In Müllerian mimicry, inedible species, usually within a taxonomic order, find it advantageous to resemble each other so as to reduce the sampling rate by predators which need to learn about the insects’ inedibility. Taxa from the toxic genus Heliconius form one of the most well known Müllerian complexes.

Chemical Defence
This is another important defence found amongst many Coleoptera and Lepidoptera species. Toxicity is usually advertised by bright stand out colours. Insects obtain their toxicity by sequestering the chemicals from the plants they eat into their own tissues however some Lepidoptera do manufacture their own toxins.
Predators that eat poisonous butterflies and moths may become sick and regurgitate or vomit violently, learning not to eat those types of species again. This is the basis of Batesian and Müllerian mimicry. A predator who has previously eaten a poisonous Lepidopteran may avoid other species with similar markings in the future, thus saving many other species as well. Some ground beetles of the Carabidae family can spray chemicals from their abdomen with great accuracy, to repel predators.
Pollination
Pollination is the process where pollen is transferred from one flower to another flower usually in the same specie so that the plant can reproduce sexually by way of natural fertilization. Most dicotyledonous flowering plants require an animal to carry out the process of transportation where as most monocotyledonous flowers rely on air currents. Insects are the chief pollinators in the animal kingdom and as such have evolved with specific traits to suit their specific plants. There is usually a mutual benefit for the insects and plants. For the pollinators it is in the form of energy rich nectar and for the plants the chance to carry its genes forth. This type of cooperation is known as mutualism.
A flower’s; traits and combinations of traits which are usually specific in attracting one or a few types of pollinators are known as pollination syndromes. These have arisen through complex plant animal adaptations. Pollinators find flowers through bright colorations, including ultraviolet light patterns, and attractant pheromones. The study of pollination by insects is known as anthecology. Bees and other social insects have a broader range of flowers to select from however some plants have still developed systems that only allow one specific ant, bee or wasp. A good example is the flowers on the Persoonia species which are Tetragonula specific. Some citrus species are able to turn the nectar off and even withdraw it if the wrong vibrations are felt thus depriving the insects of both a meal and the chance to disperse the pollen to the wrong species.
Left Native honey bee, Tetragonula sp. at hive entrance.
Right European honey bee Apis mellifera in hive- andi Mellis

Leucopogon biflorus with Tetragonula carbonaria. Upper, pair of 28 spotted ladybirds Epilachna vigintioctopunctata mating- andi Mellis

Social insects, such as termites, ants, many bees and wasps, are the most familiar species of eusocial animal. They live and work socially for the benefit of the all the members of their community caring for the brood of offspring from other individuals with a division of labour into reproductive and non-reproductive groups sometimes forming supercolonies. It is sometimes argued that the various species of honey bee are the only invertebrates to have evolved a system of abstract symbolic communication where a behavior is used to represent and convey specific information about something in the environment. The wiggle dance involves a scout bee or two wiggling in a circle dissecting the circle in the direction of the flowers relative to the position of the sun, with the extent (excitement) of the wiggling represents the type of nectar or flowers available and the length of the dance continuing until the correct number of foraging bees have gathered.
Left Termite nest of Isoptera sp. in a tree.
Right Typical termite mound of Isoptera sp. which are a common site in driers areas of Australia.

Insects which live in nests or colonies have the true capacity for fine scale spatial orientation or homing. As can be seen with bees unerringly ability to return to a single hole a few centimeters in diameter after a trip of up to several kilometers’ distance. This phenomenon is known as philopatry. Insects that hibernate have also been known to have the ability to recall a specific location up to a year later. A few insects seasonally migrate large distances between different geographic regions overwintering different locations like the Monarch butterfly.
The eusocial insects build nest, guard eggs, and provide food for offspring fulltime. Most insects, lead short lives as adults, and interactions with one another is rare except to mate or compete for mates. Even a smaller number exhibit some form of parental care, where they will guard their eggs or at times continue guard their offspring until adulthood with even fewer feeding them like the many parasitic wasps. Another simple form of parental care is to construct a nest or burrow and store provisions in it laying an egg upon those provisions. The adult does not contact the growing offspring, but it nonetheless does provide food. This sort of care is typical for most species of non eusocial bees and wasps.
Life Cycles The majority of insects hatch from eggs. The fertilization and development takes place inside the egg, enclosed by a chorion shell. In contrast to eggs of other arthropods, most insect eggs are drought resistant allowing them to be laid directly in the open under leaves and the like. This is because inside the chorion two additional membranes develop from embryonic tissue, the amnion and the serosa. This serosa secretes a cuticle; rich in chitin, that protects the embryo against desiccation.
In Schizophora however the serosa does not develop. These flies lay their eggs in moist places like rotting fruit wood or flesh. Some species of insects, like the cockroach Blaptica dubia, as well as juvenile aphids and tsetse flies, are ovoviviparous. Some other species, such as those in the genus of cockroaches known as Diploptera, are viviparous, and thus gestate inside the mother and are born alive. Some insects, like parasitic wasps, show polyembryony. Insects can be univoltine breed once a year, bivoltine breed twice a year or multivoltine where they breed more than twice a year.
Other reproductive variations have evolved and include haplodiploidy, polymorphism, paedomorphosis or peramorphosis, sexual dimorphism, parthenogenesis and more rarely hermaphroditism.
In haplodiploidy insects the sex is determined by a system where the offspring’s sex is determined by the number of sets of chromosomes an individual receives. This system is typical in bees and wasps.
Polymorphism – Polymorphism. What does it mean polymorphism? No idea love!
It is where a species may have different colour morphs or forms, as can be seen in the different colours of Tectocoris diophthalmus.

Some insects may retain phenotypes that are normally only seen in juveniles; this is called paedomorphosis.
In peramorphosis, an opposite sort of phenomenon, insects take on previously unseen traits after they have matured into adults.
Many insects display sexual dimorphism, in which males and females have notably different appearances like the females noticeably larger size difference in Dysdercus sidae.

Some insects use parthenogenesis, a process in which the female can reproduce and give birth without having the eggs fertilized by a male. Many aphids undergo a form of parthenogenesis, called cyclical parthenogenesis, in which they alternate between one or many generations of asexual and sexual reproduction. In summer, aphids are generally female and parthenogenetic while in the autumn, males may be produced for sexual reproduction. Other insects produced by parthenogenesis are bees, wasps and ants, in which they spawn males. However, overall, most individuals are female, which are produced by fertilization. The males are haploid and the females are diploid.
In a few rare cases insects display hermaphroditism, in which a given individual has both male and female reproductive organs.
Insect development undergoes two distinct forms of metamorphosis, incomplete metamorphosis and complete metamorphosis.
Insects that show incomplete metamorphosis or hemimetabolism change gradually by undergoing a series of moults. An insect moults when it outgrows its exoskeleton, which does not stretch. The process of moulting starts when the insect’s epidermis secretes a new epicuticle. After the new epicuticle is secreted, the epidermis releases a mixture of proteins that digests the endocuticle and thus detaches the old cuticle from the new hemi liquid exoskeleton. When this stage is complete, the insect makes its body swell by taking in water or air, which makes the old cuticle split along a predefined weakness in the old exocuticle. Other arthropods and crustaceans have very different processes and only moult.
Immature insects that go through incomplete metamorphosis are called nymphs or in the case of dragonflies and damselflies, naiads. Nymphs are similar in form to the adult except for the presence of wings, which do not develop until adulthood. With each moult, nymphs grow larger and become more similar in appearance to their parents.

Gulf Fritillary life cycle is an example of holometabolism or complete metamorphosis. This is where the insect changes in four stages, an egg or embryo, a larva, a pupa and the adult or imago. In these species, the egg hatches in a few minutes to 14 months to hatch and produce a larva.
The larva is generally worm, maggot or caterpillar like in form. The larva form can take one of several forms. The eruciform is a caterpillar, scarabaeiform is a grub, campodeiform is an elongated, flattened and active form, elateriform is a wire worm or vermiform is a maggot. The larva grows actively eating before it becomes a pupa.
The pupa marks a reduction in activity, and is usually sealed within a silken cocoon. There are three types of pupae: obtect, exarate or coarctate. Obtect pupae are compact, with the legs and other appendages enclosed. Exarate pupae have their legs and other appendages free and extended. Coarctate pupae develop inside the larval skin. Insects undergo considerable change known as metamorphous during the pupae stage, and emerge as adults.
Butterflies and moths are the best known examples of insects that undergo complete metamorphosis, although most insects use this life cycle. Some insects have evolved this system to hypermetamorphosis.
Complete metamorphosis is unique to a group of certain insect orders including Diptera, Lepidoptera and Hymenoptera. This form of development is exclusive to insects.
Classification
Many entomologists specialize in a single order or even a family of insects, and a number of these subspecialties are given their own informal names are usually derived from the scientific name of the group they study:
Coleopterology are beetles and they are studied by Coleopterologists.
Dipterology are flies and they are studied by Dipterologists.
Hemipterology are true bugs and they are studied by Hemipterologists.
Lepidopterology are moths and butterflies and they are studied by Lepidopterologists.
Melittology or Apiology are bees and they are studied by Melittologists or Apiologists.
Myrmecology are ants and they are studied by Myrmecologists.
Orthopterology are grasshoppers, crickets, etc. and they are studied by Orthopterologists.
Trichopterology are caddis flies and they are studied by Trichopterologists.
Vespology are social wasps and they are studied by Vespologists.
In conclusion what ever your ology is, be happy doing it.
Myriapods
Characteristics of Body structures:
Myriapods have a single pair of antennae and, in most cases, simple eyes. The mouthparts lie on the underside of the head, with an epistome and labrum forming the upper lip, and a pair of maxillae forming the lower lip. Pairs of mandibles lie inside the mouth. Myriapods breathe through spiracles that connect to a tracheal system similar to that of insects. There is a long tubular heart that extends through much of the body usually with a few, if any, blood vessels.
Malpighian tubules excrete nitrogenous waste into the digestive system, which typically consists of a simple tube. Although the ventral nerve cord has a ganglion in each segment, the brain is relatively poorly developed.
During mating, male myriapods produce a packet of sperm, or spermatophore, which they must transfer to the female externally. This process is often complex and highly developed. The female lays eggs which hatch as much shortened versions of the adults, with only a few segments and as few as three pairs of legs. The young add an additional segment and a pair of legs during each moult until they reach maturity or full adulthood.
Some of the more common Myriapod orders are:
Geophilomorpha which have 31 to 177 pairs of legs,
Lithobiomorpha which have 15 pairs of legs,
Scolopendromorpha which have 21 to 23 pairs of legs,
Scutigeromorpha which have 15 pairs of long legs.
Centipedes are carnivorous, possessing unique poison claws on the post head segment. They are usually flattened, and only have one pair of legs per segment. Centipedes are quick predators, eating any small organisms they can catch.
Millipedes are detritivores, occurring primarily in moist soils and leaf litter. They have diplosegment body parts. Each pair of trunk segments shares a single tergal plate. They usually have round bodies, and have two pairs of legs on each body segment. They move slowly and often tunnel into soil and dead leaves. Nearly all millipede species are decomposers eating dead leaves, fungi, and detritus matter. If another animal threatens them, they may curl up, and some give off smelly toxic chemicals to protect themselves.
Symphylans are detritivores, occurring primarily in moist soils and leaf litter.
Pauropoda are soft, cylindrical animals with bodies 0.5mm to 2mm in length.
Ecology
Myriapods reach their peak in moist forests as detritivorous or decomposers fulfilling an important role in decomposition of plant material. Some live in grasslands, semi arid habitats and even deserts. Most are detritivorous, with the exception of centipedes, which are chiefly nocturnal predators.
A very small percentage of species are littoral species found along the fore shores and estuaries.
Pauropodans and symphylans are small, sometimes microscopic animals that resemble centipedes superficially and live in soils.
Millipedes differ from the other groups in having their body segments fused into pairs, giving the appearance that each segment bears two pairs of legs, while the other three groups have a single pair of legs on each body segment.
Although not generally considered dangerous to humans, many millipedes produce noxious secretions; often containing benzoquinones, which in contact with the skin can cause temporary blistering and discolouration of the skin in rare cases. Large centipedes have the ability and means to inflict bites that can cause intense pain though fatalities are extremely rare.
Life Cycles: All details and figures are extracts from Brocher 1930 and Serra & (Miquel Lithobius mutabilis – Centipede) 1996.
Reproduction biology and life cycle appears to be rather consistent between the different species of Myriapods. The maximal life expectancy is around 7 years with sexual maturity being attained at the earliest age of 18 months and definitively by 2 years in the case of females. The courtships and mating behaviour involves the spinning of webs with spermatophores by males.
The eggs are laid singly with no maternal care of the brood taking place in L. mutabilis as in other Lithobiid species. The only protection for the eggs was a thin cover of fine soil particles, which is applied to the eggs by the female. Only the covered eggs and females carrying
eggs with their gonopods could be confirmed. The spherical eggs are whitish and measure 0.89mm in diameter.
Though all the eggs were kept under the same constant conditions of (16°C); the time between oviposition and hatching ranged from 21 to 284 days, most individuals hatching between 3 and 4 months in laboratories animals while those under natural conditions would hatch in 150 to 200 days. One third of the eggs hatched. Egg laying of captive animals occurred throughout the year. Other species were noted to have distinct periods for laying eggs in spring with a smaller number laid in autumn.
This different behaviour of the species and the small number of eggs laid during each oviposition period might be interpreted in correlation to habitat choice. A long egg laying period guarantees a breeding success, even if some of the eggs are destroyed through unfavourable seasonal conditions.
Females are able to conserve sperm in their seminal receptacles for up to two years personal observations on a lone female and 7 months to 12 months was not uncommon.
The high individual variability of stage duration (Fig. 5) means that new developmental and maturity varied greatly among individuals
moulting activity and stage duration depend to a large degree on the environmental conditions which were influenced mainly by food availability and low temperatures. The loss of extremities has been
simulated by Verhoeff 1915 & Scheffel 1980 under constant temperature conditions and optimal feeding where it was expected that all individuals would have a relatively similar stage duration, but
to the contrary, they varied very widely especially at the higher epimorphic stages. This is not only a phenomenon as it occurred in many other species as well.
These results using Lithobius mutabilis and other Lithobius species from around Germany suggest that the life cycles of all Centipedes at least appear to be very consistent and probably extend to the Australian species as well.
Classification
There has been much debate as to which arthropod group is most closely related to the Myriapoda. Under the Mandibulata hypothesis, Myriapoda is the sister taxon to Pancrustacea, a group comprising the Crustacea and Hexapoda. Under the Atelocerata hypothesis, Hexapoda is the closest, whereas under the Paradoxopoda hypothesis, Chelicerata is the closest. This last hypothesis, although supported by few, if any, morphological characters, is supported by a number of molecular studies. There are four classes of extant myriapods, Chilopoda the centipedes, Diplopoda, Pauropoda and Symphyla, containing a total of around 12,000 species. While each of these groups of myriapods is believed to be monophyletic, relationships among them are less certain.
Centipedes are fast moving predatory nocturnal animals that prefer cool, secluded moist locations during the day. There are around 3,000 specie world wide ranging from Nannarrup hoffmani at 12mm in length to Scolopendra gigantea of Peru at comes in at 300mm in length. Australia’s largest centipede is Ethmostigmus rubripes at a cool 160mm in length.
Ethmostigmus rubripes ranges in colour from deep blue-green-brown and olive-brown to orange-yellow. It has black bands along the body with ochre legs and antenna. The long, flattened body has 25 or 27 body segments and 21 or 23 pairs of legs. Centipedes always have an odd number of paired legs. The first pair of legs behind the head is modified to form scythe like claws which curve around its head and can inject venom into its prey. The venom is toxic to both mammals and insects, but does not appear to be strong enough to kill large animals quickly.
It is found throughout Australia except Tasmania in both dry and moist habitats, usually in sheltered places such as under logs, in leaf litter, soil, under rocks and bark in urban areas, forests, woodlands, heath, rainforests and semi arid locations.

Right – http://www.qm.qld.gov.au Centipedes
Millipedes form the class Diplopoda. Most millipedes are slower than centipedes, and feed on leaf litter and detritus. They are distinguished by the fusion of each pair of body segments into a single unit, giving the appearance of having two pairs of legs per segment. There has been around 8,000 species described world wide which estimated to be around 10mm of the actual number of specie in the world. The name “millipede” is from the Greek words mill a thousand and pedis a foot however millipedes really only have 36 and 375 legs. Millipedes always have an even number of paired legs. Illacme plenipes from the Philippines has the greatest number of legs of any animal, with 750 feet to trip over. The name pill millipedes are much shorter, and are capable of rolling up into a ball, like pillbugs.
Polydesmid millipdes is a typical Aussie having 19 to 20 body segments. They live in leaf litter or under rocks and logs. They can range in length from 1omm to 100mm in length. They don’t have eyes. When disturbed they have glands that can produce a mixture of hydrocyanide and formic acid and other chemical irritants for defense.


Symphylans are detritivores, occurring primarily in moist soils and leaf litter. They superficially resemble centipedes in appearance, but are smaller and translucent, because of their habitat being dense mulches, dark places and nocturnal. They can move rapidly through the pores between soil particles but that is where the resemblance lies.
There are around 200 species of symphylans worldwide. Many spend their lives as soil infauna with a few living arboreally. Juveniles have six pairs of legs, but, over a lifetime of several years, add an additional pair with consecutive moult so that the adult instar has twelve pairs of legs.
They are tracheal respirators breathing through the side of the head. The body comprises of 15 to 24 segments, which are protected by overlapping dorsal tergal plates. They are typically found from the surface down to a depth of about 50 cm. They consume decaying vegetation, but have been known to damage agricultural crops by consuming seeds, rootlets and root hairs in cultivated soil.
Juvenile Symphylans have six pairs of legs, but, over a lifetime of several years they add an additional pair at each moult so that the adult instar has twelve pairs of legs. Lacking eyes, their long antennae serve as sense organs.
Symphylans have several distinct features linking them to early insects like the labium or fused second maxillae, an identical number of head segments and a number of features on their legs. They are in fact a link between late Myriapods and early insects.

Pauropoda are soft, cylindrical animals with bodies 0.5mm to 2mm in length. They have neither eyesnor hearts. There are over 700 described species world wide. They are believed to be the sister group to millipedes, and have the dorsal tergites fused across pairs of segments, similar to the more complete fusion of segments seen in millipedes. The body segments have ventral tracheal, spiracular pouches forming apodemes similar to those in millipedes and Symphyla, although the trachea usually connected to these structures are absent in most species.
The juvenile first instar has three pairs of legs but, over a lifetime of several years they add an additional pair at each moult so that adult species may have nine to eleven pairs of legs.
The antennae are branching, which identifies them instantly from millipedes. No fossil pauropods have been found prior to the formation of the Baltic amber about 40 to 35 million years ago. Their head capsules show great similarities to millipedes in that they both have three pairs of mouth parts while the genital openings occur in the anterior part of the body. They also have a pupoid phase at the end of the embryonic development.

Abrus precatorius This black flower wasp was seen on the flowers prior to photographing.

Brachychiton acerifolius have Agrianome spinicollis larvae and the adult in dead branches and trunks.

Brachychiton acerifolius at times can be defoliated with the precense of leaf curl moth. Probably a Dichocrosis specie.

Rhodosphaera rhodanthema
The leaf curl moth larvae of Dichorocis species is found on the species from mid April to Late April.

Flowering occurs in September where the masses of red flowers are teaming with native hive bees like Tetragonula carbonaria.

Further Comments from Readers:
All information is included in good faith and has been thoroughly researched prior to printing. The website or the author does not warrant or guarantee the accuracy of any information on these pages, nor does the website or the author accept any responsibility for any loss arising from the use of the information found within. The views and opinions are strictly those of the author or those members who chose to actively, participate in the contents herein.
“Hi reader, it seems you use The Bible of Botany a lot. That’s great as we have great pleasure in bringing it to you! It’s a little awkward for us to ask, but our first aim is to purchase land approximately 1,600 hectares to link several parcels of N.P. into one at The Pinnacles NSW Australia, but we need your help. We’re not salespeople. We’re amateur botanists who have dedicated over 30 years to saving the environment in a practical way. We depend on donations to reach our goal. If you donate just $5, the price of your coffee this Sunday, We can help to keep the planet alive in a real way and continue to bring you regular updates and features on Australian plants all in one Botanical Bible. Any support is greatly appreciated. Thank you.”
In the spirit of reconciliation we acknowledge the Bundjalung, Gumbaynggirr and Yaegl and all aboriginal nations throughout Australia and their connections to land, sea and community. We pay our respect to their Elders past, present and future for the pleasures we have gained.