Concepts:
After almost 200 years Charles Darwin’s published theories on “Evolution” and “The Origin of a Species” still remains controversial. It has had such an impact on society especially in the west between believers in Biblical Creation and those who ascribe to Darwin’s theories.
The major point of Darwin’s theory is that species are created by the process of natural selection. Organisms evolve through a process of “survival of those which have the traits most adapted to environmental changes,” where the organisms best adapted to environmental changes within their niche survive and those that are not so well adapted become extinct. Eventually, change can occur to the extent that a new species exists that is different from its ancestor species.
This is important for biologists because it explains what is now the most commonly accepted theory of how speciation occurs.

Despite the advances that humans have made, no other living creature since the incept of living cells has had such a profound alteration on Earth’s environment, yet natural selection continues to work on our species as it is on all species that survive the present extinction rate.
After almost 200 years Charles Darwin’s published theories on “Evolution” and “The Origin of a Species” still remains controversial. It has had such an impact on society especially in the west between believers in Biblical Creation and those who ascribe to Darwin’s theories.
The major point of Darwin’s theory is that species are created by the process of natural selection. Organisms evolve through a process of “survival of those which have the traits most adapted to environmental changes,” where the organisms best adapted to environmental changes within their niche survive and those that are not so well adapted become extinct. Eventually, change can occur to the extent that a new species exists that is different from its ancestor species.
This is important for biologists because it explains what is now the most commonly accepted theory of how speciation occurs.

This pretty face wallaby; Macropus parryi, is perfectly suited to drier wallums which are overgrown with long grasses and reed like plants.
Evolution can be explained as slow on going process by which modern organisms have descended from ancient organisms.
The theory of evolution is now well documented and supported with both living and fossilized past occurrences naturally laid before us over the millenniums.
Darwin’s initial observations were that of those organisms he had collected, they were all perfectly suited to their environmental niche. More over, every species had a combination of physical characteristics and behaviours that assisted to survive and adapt. Catching food, deterring enemies, with stand local variants in habitat conditions and the ability to reproduce successfully.


In addition Darwin collected fossils on his travels and during quiet periods at sea especially on the later part of the journey began putting the pieces together. His keen observational powers were beginning to unravel the existence of lines and linkages to extinct organisms and those in other regions.
Some of Darwin’s fossils were unusual but it was the fossils which were similar to modern day organisms that held his desire to expound the hidden secrets they held.
Darwin was puzzled and made more curious by the stark resemblance of many living organisms to those of organisms that were now extinct.
These similarities were too consistent and raised such issues as to why did they become extinct if they were in fact so similar? We now know that Fossil diversity like the present day diversity depicts a changing environment where only those adapted to change can and will survive.

By collecting plant and animal specimens from many different environments Darwin began to fit together a surprising jigsaw puzzle that contradicted past theories of the origin of humans, animals and plants. For the first time someone was detecting small differences in the same organism from different places which experienced small differences in environmental habitats.
The Galapagos Islands start to unravel the mysteries. The Galapagos Islands consist of 15 small volcanic Islands which lie 970 kilometres west of Ecuador and slightly below the equator. Because of their different sizes each island had a slightly different landscape which in return changed the climatic and microclimate on the individual island.
It was here that Darwin had his first strong evidence that environmental conditions or at least the isolation of a species with different environmental conditions give rise to different genes adapting better to different environmental conditions.
Darwin noticed that while the animals were different they were essentially similar to those animals found on the mainland of the South American continent.
It wasn’t until later in the journey that Darwin realised that the animals he had observed on the Galapagos Islands were also different to each other and he could identify a particular animal with a particular Island in the group or the food niche it occupied. These included the Land Tortoises, the Marine Iguanas and the Darwin Finches which he is well noted for.

The beaks, song and colour of the 14 species of finches vary accordingly to the diets of the birds which have changed since the arrival of their common ancestor about 2.5 million to 3 million years ago on the 4.7 million year old Islands.
Isla Espanola Island Santa Cruz Island Isabela Island
The Marine Iguanas on the different Islands are unique in that they are the worlds only marine Lizard, have their own distinct size, shape and color.

Darwin’s curiosity led him to think that maybe the different groups of animals on the different Islands once had common ancestors and in fact evolved after they were isolated from their ancestors arrival and isolated from one another.
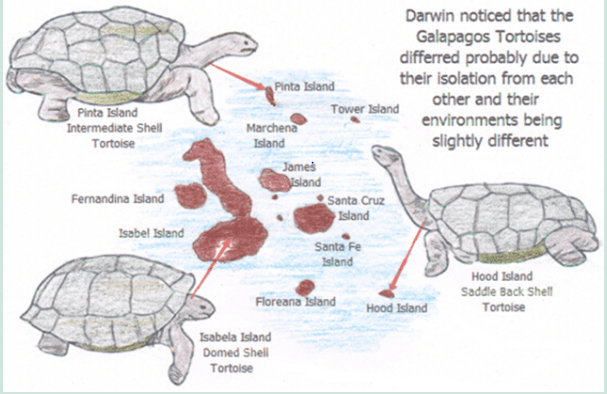

Darwin’s curiosity led him to think that maybe the different groups of animals on the different Islands once had common ancestors and in fact evolved after they were isolated from their ancestors arrival and isolated from one another.
Darwin began to challenge his own beliefs; following the voyage of the Beagle, as he could not believe the Earth was stable and unable of change. Many scholars especially those of the church believed that the world was created just a few thousand years prior to his day and that all living things were divine creatures of a divine intervention. The evidence he had collected from around the world did not fit into any previous thoughts, education and religious studies he had undertaken. James Hutton and Charles Lyell two emminent geologists spark the fire to reshape Darwin’s thinking immeasurably.
James Hutton after some 25 years of research he published his “Theory of the Earth,” in an “Investigation of the Laws observable in the Composition, Dissolution, and Restoration of Land upon the Globe.” Basically he theorized that the solid parts of the present land appear in general, to have been composed of the productions of the sea, and of other materials similar to those now found upon the shores thus he found reason to conclude:
1st: That the land on which we rest is not simple and original, but that it is a composition, and had been formed by the operation of the second causes.
2nd: That before the present land was made, there had subsisted a world composed of sea and land, in which there were tides and currents, with such operations at the bottom of the sea as they are now taking place.
3rd: That while the present land was forming at the bottom of the ocean, the former land maintained plants and animals; at least the sea was then inhabited by animals, in a similar manner as it is at present.
Thus he concluded, that the greater part of our land, if not the whole had been produced by operations natural to this globe; but that in order to make this land a permanent body, resisting the operations of the waters, two things had been required;
1st: The consolidation of masses formed by collections of loose or incoherent materials;
2nd: The elevation of those consolidated masses from the bottom of the sea, the place where they were collected, to the stations in which they now remain above the level of the ocean.
James Hutton’s search for evidence:
At Glen Tilt in the Cairngorm mountains in the central Scottish Highlands in 1785; James Hutton found granite penetrating metamorphic schists, in a manner which indicated that the granite had been molten at the time of formation. This showed to him that the granite formed from cooling of molten rock, not precipitation out of water as others at the time believed, and that the granite must be younger than the surrounding schists.

He located similar penetrations of volcanic rock through sedimentary rocks near the coast the centre of Edinburgh, at Salisbury Crags, adjoining Arthur’s Seat. Later in 1786, he recovered more evidence in Galloway and on the Isle of Arran in 1787.

The existence of angular unconformities had been noted by Nicolas Steno and by French geologists including Horace-Bénédict de Saussure, who interpreted them in terms of Neptunism as “primary formations”. Hutton wanted to examine such formations himself to see “particular marks” of the relationship between the rock layers. On the 1787 trip to the Isle of Arran he found his first example of Hutton’s Unconformity to the north of Newton Point near Lochranza, but the limited view meant that the condition of the underlying strata was not clear enough for him, and he incorrectly thought that the strata were conformable at a depth below the exposed outcrop.

Later in 1787 Hutton noted what is now known as the Hutton or “Great” Unconformity at Inchbonny, Jedburgh, in layers of sedimentary rock. As shown in the photograph below, layers of greywacke can be seen in the lower right corner and again on the central right of the cliff face are tilted almost vertically. The intervening layer of conglomerate rock lies almost horizontally over stratum of old red sandstone. He later wrote of how he “rejoiced at my good fortune in stumbling upon an object so interesting in the natural history of the earth, and which I had been long looking for in vain.” That year, he found the same sequence near Teviotdale.

In the spring of 1788 he and John Playfair researched the area near the Berwickshire coast and found more examples of this sequence in the valleys of the Tour and Pease Burns near Cockburnspath. On the cliffs below St. Helens; just to the east at Siccar Point, found what Hutton called “A beautiful picture of this junction washed bare by the sea.” Continuing along the coast, they made more discoveries including sections of the vertical beds showing strong ripple marks which gave Hutton “great satisfaction” as a confirmation of his supposition that these beds had also been laid horizontally in water. He also found conglomerate at altitudes that demonstrated the extent of erosion of the strata.
Hutton reasoned that there must have been innumerable cycles, each involving deposition on the seabed, uplift with tilting and erosion then again returned beneath the sea for further layers to be deposited. On the belief that this was due to the same geological forces operating in the past as the very slow geological forces seen operating at the present day, the thicknesses of exposed rock layers implied to him enormous stretches of time.
Charles Lyell had private means, and earned further income as an author. He came from a prosperous family, worked briefly as a lawyer in the 1820s, and held the post of Professor of Geology at King’s College London in the 1830s. From 1830 onward his books provided both income and fame. His three major books were works which were continually in progress. All three books went through multiple editions during his lifetime, although many of his friends; including Charles Darwin, thought the first edition of the Principles was the best written. Lyell used each edition to incorporate additional material, rearrange existing material, and revisit old conclusions in light of new evidence.

Principles of Geology was Lyell’s first book, was also his most famous, most influential, and most important. First published in three volumes in 1830–33, it established Lyell’s credentials as an important geological theorist.
The basic argument in Principles was that “The present is the key to the past.” His concept states “What has happened in the past is happening now and will happen in the future thereafter. Geological remains from the distant past can, and should, be explained by reference to geological processes now in operation and thus are directly observable.”
Lyell’s interpretation of geologic change is the steady accumulation of minute changes over enormously long spans of time was a powerful influence on the young Charles Darwin. Lyell asked Robert FitzRoy, captain of HMS Beagle, to search for erratic boulders on the survey voyage of the Beagle, and just before it set out Fitzroy gave Darwin Volume 1 of the first edition of Lyell’s Principles. When the HMS Beagle made its first stop ashore at St Jago, Darwin found rock formations which seen “through Lyell’s eyes” gave him a revolutionary insight into the geological history of the island, an insight he applied throughout his travels.
While in South America Darwin received Volume 2 in which considered the ideas of Lamarck in some detail. In geology Darwin was very much Lyell’s disciple, and brought back observations and his own original theories, including ideas about the formation of atolls, which supported Lyell’s uniformitarianism. On the return of the HMS Beagle on October 1836, Lyell invited Darwin to dinner and from then on they were close friends. Although Darwin discussed evolutionary ideas with him from 1842, Lyell continued to reject evolution in each of the first nine editions of the Principles. Despite his personal views at the time he encouraged Darwin to publish his findings, and following the 1859 publication of On the Origin of Species, Lyell finally offered a tepid endorsement of evolution in the tenth edition of Principles.
Elements of Geology
Geological Evidences of the Antiquity of Man brought together Lyell’s views on three key themes from the geology of the Quaternary Period of the Earth’s history – glaciers, evolution, and the age of the human race.
Uniformitarian:
Principles of Geology was “An attempt to explain the former changes of the Earth’s surface by reference to causes now in operation”, and this explains Lyell’s impact on science. He drew his explanations from field studies conducted directly before he went to work on the founding geology text. He was, along with the earlier John Playfair, who was the major advocate of James Hutton’s idea of uniformitarianism, that the earth was shaped entirely by slow moving forces still in operation today, acting over a very long period of time. This was in contrast to catastrophism, a geologic idea of abrupt changes, which had been adapted in England to support belief in Noah’s flood.
Stratigraphy:
Lyell’s most important specific work was in the field of stratigraphy. From May 1828, until February 1829, he travelled with Roderick Impey Murchison (1792–1871) to the south of France (Auvergne volcanic district) and to Italy. In these areas he concluded that the recent strata (rock layers) could be categorised according to the number and proportion of marine shells encased within. Based on this he proposed dividing the Tertiary period into three parts, which he named the Pliocene, Miocene, and Eocene.
He also renamed the traditional Primary, Secondary and Tertiary periods which are now known as the Paleozoic, Mesozoic and Cenozoic Eras.
His observational methods and general analytical framework have stood the test of time and remain in use today as foundational principles in geology.
Evolution
Lyell first received a copy of one of lamarck’s books from Mantell in 1827, when he was on circuit. Though he disagreed with most the content it is known that he did acknowledge that the Earth is much older than people had previously thought.
His final resolution was if the earth itself could change through rivers, glaciers, Volcanoes and earthquakes then why couldn’t nature change to adapt to the changing conditions.
Lamarck’s Theory of Evolution influence Darwins thought:
Lamark proposed a theory that by the selective use or disuse of an organ/s by organisms, the organisms acquired or lost certain traits during their lifetime. These traits could be passed onto their offspring and over a period of time would lead to a permanent change/s within the organisms.
It is fascinating to remark on the differences between the theories of Lamarck and Darwin. A superficial glance does not reveal any great difference between the two theories but it is apparent that Lamarck refers to the individual organism being able to change while Darwin refers to the evolution acting on whole populations.
Lamarck’s theory was that a species underwent changes in response to changes in their environment. One example referred by him was that of Giraffes. As a response to trees in their environment growing taller, giraffes responded to the change by growing longer necks so that they could continue to eat the leaves they ate before.

His second theory was that this change was permanent and on going for as long as the new environmental conditions continued to apply. This is where nature chose the best possible solution and organisms responded accordingly.
Darwin and Wallace’s theory was one of natural selection and survival of the fittest. As the environment underwent changes, the species affected by these changes underwent changes in response to the changes in the environment. It appears that there is not much dissimilarity between the two theories proposed by Lamarck and that of Darwin but what does make a difference is that Darwin proposed a mechanism whereby such changes were effective. Changes in species were due to procreation or breeding. Those species that spawned changes that adapted to the new conditions, survived while those that didn’t died out.
With this relatively rudimentary distinction between the two theories, the vast differences between the two become apparent. Lamarck envisioned, with no experimental observation to back his hypothesis, changes taking place singly and automatically in response to changes in the environment, corresponding to a mechanical working of nature; the environment underwent a change and nature underwent a change. By contrast, Darwin saw infinite changes taking place in nature evoking infinite responses in organisms where only with the fittest or most adaptable would survive.
The differences between the theories of Lamarck and Darwin, illustrates vividly the difference between philosophical musings, even though they might be surprisingly intuitive and close to the heart of the problem, and a rigorous investigation based on observation supported by consistent data.
Yet what we often forget, in our bedazzlement at Darwin’s brilliance, is that Lamarck’s idea that organisms respond to changes in their environment was transcendental and instrumental in the formulation of Darwin’s theory of evolution.
The English Economist Thomas Malthus observed in 1778 that babies were being born faster than people were dying and stated that if the population continued to grow the world would be overrun by people.
He also stated that war, famine and disease would eventually limit the human expansion.
Darwin with reasoning realized that the same applied to plants and animals.
Alfred Russel Wallace and Charles Darwin:

Much of what has been written below are extracts from: Wallace’s autobography. – www.alfredwallace.org,nhm.ac.uk/nature-online/evolution, www.reasons.org/articles/alfred-russel-wallace-and-intelligent-evolution, coupled with the authors views on Wallace.
Lyell and Hooker were instrumental in arranging the peaceful co-publication of the theory of natural selection by Darwin and Alfred Russel Wallace in 1858: Each had arrived at the theory independently. Lyell’s data on stratigraphy were important because Darwin thought that populations of an organism changed slowly, requiring “geologic time.”
Russel Wallace conducted most his research on Ternate Island in Indonesia and it was from there that he sent his famous paper dated February 1858, addressed to Darwin. It was entitled “On the Tendency of Varieties to Depart Indefinitely from the Original Type.” Wallace’s paper was a beautifully written scientific treatise, which defined the mechanism of the origin of new species. The paper made mention of the idea of the “Survival of the fittest” as a result of natural selection.
The paper was meant for publication in a prestigious journal but instead was scheduled for presentation at the forthcoming meeting of the Linnean Society in London.
When Darwin received Wallace’s paper he was horrified to learn that it dwelt precisely on the same theory that he had agonized and modified time and time again. Darwin had spent more than 20 years assembling his field notes and information and making refinements that supported his theory on the “Origin of species”. He agonized over the fact that his lifetime work now faced imminent desolation.
Charles Lyell, Charles Darwin’s friend arranged that the independent works of the two scientists be presented simultaneously. The papers of Darwin and Wallace were read on July 1, 1858 at the Linnean Society meeting in London. Since then, both scientists have been credited with being the independent co-discoverers of the theory of evolution based on natural selection.
Wallace began his career as a travelling naturalist already believing in the transmutation of species. The concept had been advocated by Jean-Baptiste Lamarck, Geoffroy Saint-Hilaire, Erasmus Darwin, and Robert Grant, among others. It was widely discussed, but not generally accepted by leading naturalists, and was considered to have radical, even revolutionary connotations.
Prominent anatomists and geologists including Georges Cuvier, Richard Owen, Adam Sedgwick, and Charles Lyell attacked were strong advocates against evolution on the grounds of creation. It is believed that Wallace accepted the idea of the transmutation of species in part because he was always inclined to favour more radical ideas in politics as a socialist. As a young man Wallace had seen poverty and the intrusive and oppressive effects of the powerful elite over farmers in the implementation of the Enclosure Acts in rural Wales. He also witnessed first hand working class life and the harsh conditions his brother endured as an apprentice in London. Those experiences affected Wallace and stayed with him all his life. But Wallace’s brand of socialism was based upon social cooperation and was wary of Marxist militancy and government coercion. He showed a preference for individual liberty over government mandates and coercive policy enactments. There is a libertarian strain in much of Wallace’s political philosophy. Nonetheless, the broader implication was that of his belief that science especially the natural sciences should be recognized and elevated to the highest order.
Religion and science and because he was unusually open to marginal, even fringe, ideas in science opened up the ability to reason and think beyond the scope of most people. This extended to wanting to use the human brain as a tool in science. Wallace was almost entirely self taught and as such was without many influential friends. These are probably the most important single reason why he was attacked more than Darwin and given far less prominence by his peers. Nevertheless his works still stand shoulder to shoulder with Darwin on evolution amongst biologists and geologists.
Wallace’s vision coupled with his interest in biogeography resulted in him being one of the first prominent scientists to raise concerns over the environmental impact of human activity.
Wallace was also profoundly influenced by Robert Chambers’ work Vestiges of the Natural History of Creation, a highly controversial work of popular science published anonymously in 1844 that advocated an evolutionary origin for the solar system, the earth, and living things.
Wallace wrote to Henry Bates in 1845:
“I have a rather more favourable opinion of the ‘Vestiges’ than you appear to have. I do not consider it a hasty generalization, but rather as an ingenious hypothesis strongly supported by some striking facts and analogies, but which remains to be proven by more facts and the additional light which more research may throw upon the problem. It furnishes a subject for every student of nature to attend to; every fact he observes will make either for or against it, and it thus serves both as an incitement to the collection of facts, and an object to which they can be applied when collected.”
In 1847, he wrote to Bates:
“I should like to take some one family; of beetles, to study thoroughly, principally with a view to the theory of the origin of species. By that means I am strongly of opinion that some definite results might be arrived at.”
Wallace deliberately planned his field work to test his hypothesis but was open minded to change tact if circumstances dictated. Under an evolutionary scenario closely related species should inhabit neighbouring territories. During his work in the Amazon basin, he came to realize that geographical barriers—such as the Amazon and its major tributaries—often separated the ranges of closely allied species, and he included these observations in his 1853 paper “On the Monkeys of the Amazon”. Near the end of the paper he asks the question “Are very closely allied species ever separated by a wide interval of country?”
In February 1855, while working in Sarawak on the island of Borneo, Wallace wrote “On the Law which has Regulated the Introduction of New Species”, a paper which was published in the Annals and Magazine of Natural History in September 1855. In this paper, he discussed observations regarding the geographic and geologic distribution of both living and fossil species. It would later become known as biogeography. His conclusion he said “Every species has come into existence coincident both in space and time with a closely allied species” It became known as the “Sarawak Law”. Wallace thus answered the question he had posed in his earlier paper on the monkeys of the Amazon River basin.
It made no mention of any possible connection to evolution however this paper foreshadowed the momentous paper he would write three years later.
The paper shook Charles Lyell’s belief that species were immutable. Although his friend Charles Darwin had written to him in 1842 expressing support for transmutation, Lyell had continued to be strongly opposed to the idea. Around the start of 1856, he told Darwin about Wallace’s paper, as did Edward Blyth who thought it was “Good! Upon the whole! … Wallace has, I think put the matter well; and according to his theory the various domestic races of animals have been fairly developed into species.” Despite this hint, Darwin mistook Wallace’s conclusion for the progressive creationism of the time and wrote that it was “nothing very new ….. Uses my simile of tree [but] it seems all creation with him.” Lyell was more impressed, and opened a notebook on species, in which he grappled with the consequences, particularly for human ancestry”. Darwin had already shown his theory to their mutual friend Joseph Hooker and now, for the first time, he spelt out the full details of natural selection to Lyell. Although Lyell could not agree, he urged Darwin to publish to establish priority. Darwin declined at first, then began writing up a species sketch of his continuing work in May 1856.
Natural selection and Darwin:
By February 1858, Wallace was totally convinced that his biogeographical research in the Malay Archipelago that the reality of evolution was entirely factual. He later wrote in his autobiography:
“The problem then was not only how and why do species change, but how and why do they change into new and well defined species, distinguished from each other in so many ways; why and how they become so exactly adapted to distinct modes of life; and why do all the intermediate grades die out – as geology shows they have died out and leave only clearly defined and well marked species, genera, and higher groups of animals?”
According to his autobiography, it was while he was in bed with a fever that Wallace thought about Thomas Malthus’s idea of positive checks on human population growth and came up with the idea of natural selection. Wallace said in his autobiography that he was on the island of Ternate at the time; but historians have questioned this, saying that on the basis of the journal he kept at the time, he was on the island of Gilolo. From 1858 to 1861 he rented a house on Ternate from the Dutchman M.D. van Renesse van Duivenbode. He used this house as a base camp for expeditions to other islands such as Gilolo.
Wallace describes how he discovered natural selection as follows:
“It then occurred to me that these causes or their equivalents are continually acting in the case of animals also; and as animals usually breed much more quickly than does mankind, the destruction every year from these causes must be enormous in order to keep down the numbers of each species, since evidently they do not increase regularly from year to year, as otherwise the world would long ago have been crowded with those that breed most quickly. Vaguely thinking over the enormous and constant destruction which this implied, it occurred to me to ask the question, why do some die and some live? And the answer was clearly, on the whole the best fitted live … and considering the amount of individual variation that my experience as a collector had shown me to exist, then it followed that all the changes necessary for the adaptation of the species to the changing conditions would be brought about … In this way every part of an animals organization could be modified exactly as required, and in the very process of this modification the unmodified would die out, and thus the definite characters and the clear isolation of each new species would be explained.”
Wallace had once briefly met Darwin, and was one of the correspondents whose observations Darwin used to support his own theories. Although Wallace’s first letter to Darwin has been lost, Wallace carefully kept the letters he received. In the first letter, dated 1 May 1857, Darwin commented that Wallace’s letter of 10 October which he had recently received, as well as Wallace’s paper “On the Law which has regulated the Introduction of New Species” of 1855, depicted that the two men were clearly both thinking alike and to some extent reaching similar conclusions, and said that he was preparing his own work for publication in about two years time.
The second letter, dated 22 December 1857, Darwin depicted how glad he was that Wallace was theorizing about distribution, adding that “without speculation there is no good and original observation” while commenting that “I believe I go much further than you.”
Wallace trusted Darwin’s opinion on the matter and sent him his February 1858 essay, “On the Tendency of Varieties to Depart Indefinitely From the Original Type”, with the request that Darwin would review it and pass it on to Charles Lyell if he thought it worthwhile. Although Wallace had sent several articles for journal publication during his travels through the Malay archipelago, the Ternate essay was in a private letter.
On 18 June 1858, Darwin received the essay from Wallace. While Wallace’s essay obviously did not employ Darwin’s term “natural selection”, it did outline the mechanics of an evolutionary divergence of species from similar ones due to environmental pressures. In this sense, it was very similar to the theory that Darwin had worked on for twenty years, but had yet to publish. Darwin sent the manuscript to Charles Lyell with a letter saying “he could not have made a better short abstract! Even his terms now stand as heads of my chapters … he does not say he wishes me to publish, but I shall, of course, at once write and offer to send to any journal.
Distraught about the illness of his baby son, Darwin put the problem to Charles Lyell and Joseph Hooker, who decided to publish the essay in a joint presentation together with unpublished writings which highlighted Darwin’s priority. Wallace had not asked for publication of his essay, and indeed, doing so probably contravened the copyright law of the time. Wallace’s essay was presented to the Linnean Society of London on 1 July 1858, along with excerpts from an essay which Darwin had disclosed privately to Hooker in 1847 and a letter Darwin had written to Asa Gray in 1857.
Communication with Wallace in Malay was impossible without months of delay, so he was not part of this rapid publication. Fortunately, Wallace accepted the arrangement after the fact, happy that he had been included at all, and never expressed public or private bitterness. Darwin’s social and scientific status was far greater than Wallace’s, and it was unlikely that, without Darwin, Wallace’s views on evolution would have been taken seriously. Lyell and Hooker’s arrangement relegated Wallace to the position of co-discoverer, and he was not the social equal of Darwin or the other prominent British natural scientists. However, the joint reading of their papers on natural selection associated Wallace with the more famous Darwin. This, combined with Darwin’s, as well as Hooker’s and Lyell’s advocacy on his behalf, would give Wallace greater access to the highest levels of the scientific community. The reaction to the reading was muted, with the president of the Linnean Society remarking in May 1859 that the year had not been marked by any striking discoveries. Darwin’s publication of “On the Origin of Species” later in 1859, its significance became apparent. When Wallace returned to the UK, he met Darwin. Although some of Wallace’s iconoclastic opinions in the ensuing years would test Darwin’s patience, they remained on friendly terms for the rest of Darwin’s life.
Over the years, a few people have questioned this version of events. In the early 1980s, two books, one written by Arnold Brackman and another by John Langdon Brooks, even suggested not only that there had been a conspiracy to rob Wallace of his proper credit, but that Darwin had actually stolen a key idea from Wallace to finish his own theory. These claims have been examined in detail by a number of scholars who have not found them to be convincing. Research into shipping schedules has shown that, contrary to these accusations, Wallace’s letter could not have been delivered earlier than the date shown in Darwin’s letter to Lyell.
Defence of Darwin and his ideas:
After the publication of Darwin’s On the Origin of Species, Wallace became one of its staunchest defenders on his return to England in 1862. In one incident in 1863 that particularly pleased Darwin, Wallace published the short paper “Remarks on the Rev. S. Haughton’s Paper on the Bee’s Cell, And on the Origin of Species” in order to rebuke a paper by a professor of geology at the University of Dublin that had sharply criticised Darwin’s comments in the Origin on how hexagonal honey bee cells could have evolved through natural selection.
An even lengthier defence of Darwin’s work was “Creation by Law”, a review Wallace wrote in 1867 for The Quarterly Journal of Science of the book The Reign of Law, which had been written by George Campbell, the 8th Duke of Argyll, as a refutation of natural selection.
After an 1870 meeting of the British Association, Wallace wrote to Darwin complaining that there were “no opponents left who know anything of natural history, so that there are none of the good discussions we used to have.”
Differences between Darwin’s and Wallace’s ideas on natural selection:
Historians of science have noted that, while Darwin considered the ideas in Wallace’s paper to be essentially the same as his own, there were differences. Darwin emphasized competition between individuals of the same species to survive and reproduce, whereas Wallace emphasized environmental pressures on varieties and species forced them to become adapted to their local conditions or die, which lead populations in different habitats to diverge. Some historians, including Peter J. Bowler, have suggested the possibility that in the paper he mailed to Darwin, Wallace was not discussing selection of individual variations at all but rather group selection.
Others have noted that another difference was that Wallace appeared to have envisioned natural selection as a kind of feedback mechanism keeping species and varieties adapted to their environment. They point to a largely overlooked passage of Wallace’s famous 1858 paper:
“The action of this principle is exactly like that of the centrifugal governor of the steam engine, which checks and corrects any irregularities almost before they become evident; and in like manner no unbalanced deficiency in the animal kingdom can ever reach any conspicuous magnitude, because it would make itself felt at the very first step, by rendering existence difficult and extinction almost sure soon to follow.”
The cybernetician and anthropologist Gregory Bateson observed in the 1970s that, though writing it only as an example, “Wallace had probably delivered the most powerful thing that had been said in the 19th Century.” Bateson revisited the topic in his 1979 book Mind and Nature: A Necessary Unity, and other scholars have continued to explore the connection between natural selection and systems theory.
Warning colouration and sexual selection:
In 1867, Darwin wrote to Wallace about a problem he was having understanding how some caterpillars could have evolved conspicuous colour schemes. Darwin had come to believe that sexual selection, an agency to which Wallace did not pay the same significance to as Darwin did to explain many conspicuous animal colour schemes. However, Darwin realized that this could not apply to caterpillars. Wallace responded that he and Henry Bates had observed that many of the most spectacular butterflies had a peculiar odour and taste, and that he had been told by John Jenner Weir that birds would not eat a certain kind of common white moth because they found it unpalatable. “Now, as the white moth is as conspicuous at dusk as a coloured caterpillar in the daylight”, Wallace wrote back to Darwin that it seemed likely that the conspicuous colour scheme served as a warning to predators and thus could have evolved through natural selection. Darwin was impressed by the idea and at a subsequent meeting of the Entomological Society, Wallace asked for any evidence anyone might have on the topic.
In 1869, Weir published data from experiments and observations involving brightly coloured caterpillars that supported Wallace’s idea. Warning colouration was one of a number of contributions Wallace made in the area of the evolution of animal colouration in general and the concept of protective colouration in particular.
A lifelong disagreement Wallace had with Darwin was over the importance of sexual selection. In his 1878 book “Tropical Nature and Other Essays,” he wrote extensively on the colouration of animals and plants and proposed alternative explanations for a number of cases Darwin had attributed to sexual selection.
He revisited the topic at length in his 1889 book Darwinism. In 1890, he wrote a critical review in Nature of his friend Edward Bagnall Poulton’s “The Colours of Animals” which supported Darwin on sexual selection, attacking especially Poulton’s claims on the “æsthetic preferences of the insect world”.
The Wallace effect on hybrids between closely related species:
In 1889, Wallace wrote the book “Darwinism,” which explained and defended natural selection. In it, he proposed the hypothesis that natural selection could drive the reproductive isolation of two varieties by encouraging the development of barriers against hybridization. Thus it might contribute to the development of new species. He suggested the following scenario. When two populations of a species had diverged beyond a certain point, each adapted to particular conditions, hybrid offspring would be less well-adapted than either parent form and, at that point, natural selection will tend to eliminate the hybrids. Furthermore, under such conditions, natural selection would favour the development of barriers to hybridization, as individuals that avoided hybrid matings would tend to have more fit offspring, and thus contribute to the reproductive isolation of the two incipient species. This idea came to be known as the Wallace effect.
Wallace had suggested to Darwin that natural selection could play a role in preventing hybridization in private correspondence as early as 1868, but had not worked it out to this level of detail. It continues to be a topic of research in evolutionary biology today, with both computer simulation and empirical results supporting its validity.
Application of theory to humans, and role of teleology in evolution.
An illustration from the chapter on the application of natural selection to humans in Wallace’s 1889 book Darwinism shows a chimpanzee.
In 1864, Wallace published a paper, “The Origin of Human Races and the Antiquity of Man Deduced from the Theory of ‘Natural Selection'”, applying the theory to humankind. Darwin had not yet publicly addressed the subject, although Thomas Huxley had in “Evidence as to Man’s Place in Nature.” He explained the apparent stability of the human stock by pointing to the vast gap in cranial capacities between humans and the great apes. Unlike some other Darwinists, including Darwin himself, he did not “regard modern primitives as almost filling the gap between man and ape”. He saw the evolution of humans in two stages: achieving a bipedal posture freeing the hands to carry out the dictates of the brain, and the “recognition of the human brain as a totally new factor in the history of life. Wallace was apparently the first evolutionist to recognize clearly that … with the emergence of that bodily specialization which constitutes the human brain, bodily specialization itself might be said to be outmoded.” For this paper he won Darwin’s praise.
Shortly afterwards, Wallace became a spiritualist. At about the same time, he began to maintain that natural selection cannot account for mathematical, artistic, or musical genius, as well as metaphysical musings, and wit and humour. He eventually said that something in “the unseen universe of Spirit” had interceded at least three times in history. The first was the creation of life from inorganic matter. The second was the introduction of consciousness in the higher animals. And the third was the generation of the higher mental faculties in humankind. He also believed that the raison d’être of the universe was the development of the human spirit. These views greatly disturbed Darwin, who argued that spiritual appeals were not necessary and that sexual selection could easily explain apparently non-adaptive mental phenomena. While some historians have concluded that Wallace’s belief that natural selection was insufficient to explain the development of consciousness and the human mind was directly caused by his adoption of spiritualism, while other Wallace scholars have disagreed, and some maintain that Wallace never believed natural selection applied to those areas. Reaction to Wallace’s ideas on this topic among leading naturalists at the time varied. Charles Lyell endorsed Wallace’s views on human evolution rather than Darwin’s. Wallace’s belief that human consciousness could not be entirely a product of purely material causes was shared by a number of prominent intellectuals in the late 19th and early 20th centuries. However, many, including Huxley, Hooker, and Darwin himself, were critical of Wallace. As the historian of science Michael Shermer has stated, Wallace’s views in this area were at odds with two major tenets of the emerging Darwinian philosophy, which were that evolution was not teleological predetermined and that it was not peculiar to humans alone.
Much later in his life Wallace returned to these themes, that evolution suggested that the universe might have a purpose and that certain aspects of living organisms might not be explainable in terms of purely materialistic processes, in a 1909 magazine article entitled “The World of Life,” which he later expanded into a book of the same name; a work that Shermer said anticipated some ideas about design in nature and directed evolution that would arise from various religious traditions throughout the 20th century.
Assessment of Wallace’s role in history of evolutionary theory:
In many accounts of the development of evolutionary theory, Wallace is mentioned only in passing as simply being the stimulus to the publication of Darwin’s own theory. In reality, Wallace developed his own distinct evolutionary views which diverged from Darwin’s and was considered by many especially Darwin to be a leading thinker on evolution in his day, whose ideas could not be ignored.
Darwin and Wallace exchanged knowledge through both private correspondence and published works and stimulated each other’s ideas and theories over an extended period. Wallace is the most-cited naturalist in Darwin’s “Descent of Man,” often in strong disagreement. Wallace remained an ardent defender of natural selection for the rest of his life. By the 1880s, evolution was widely accepted in scientific circles. Wallace and August Weismann were nearly alone among prominent biologists in believing that natural selection was the major driving force behind it.
In 1889, Wallace published the book “Darwinism” as a response to the scientific critics of natural selection. Of all Wallace’s books, it is the most cited by scholarly publications to this day.
Biogeography and ecology:
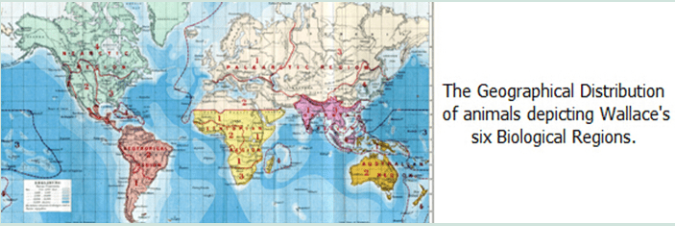
In 1872, at the urging of many of his friends, including Darwin, Philip Sclater, and Alfred Newton, Wallace began research for a general review of the geographic distribution of animals. He was unable to make much progress initially, in part because classification systems for many types of animals were in flux at the time. He resumed the work in earnest in 1874 after the publication of a number of new works on classification. Extending the system developed by Sclater for birds – which divided the earth into six separate geographic regions for describing species distribution – to cover mammals, reptiles and insects as well? Wallace created the basis for the zoogeographic regions still in use today. He discussed all of the factors then known to influence the current and past geographic distribution of animals within each geographical region. These included the effects of the appearance and disappearance of land bridges. Like those that presently connect North and South America and the Wallace line between Australia and Asia which are direct effects of periods of increased glacial activities. He provided maps that displayed factors, such as elevation of mountains, depths of oceans, and the character of regional vegetation, that affected the distribution of animals. He also summarized all the known families and genera of the higher animals and listed their known geographic distributions.
The text was organized so that it would be easy for a traveller to learn what animals could be found in a particular location. The resulting two-volume work, “The Geographical Distribution of Animals,” was published in 1876 and would serve as the definitive text on zoogeography for the next 80 years.
In “The Geographical Distribution of Animals,” Wallace did not confine himself to the biogeography of living species, but extended it to included evidence from the fossil record to discuss the processes of evolution and migration that had led to the geographical distribution of modern animal species. For example, he discussed how fossil evidence showed that tapirs had originated in the Northern Hemisphere, migrating between North America and Eurasia and then, much more recently, to South America after which the northern species became extinct, leaving the modern distribution of two isolated groups of tapir species in South America and Southeast Asia.
Wallace was very aware of, and interested in, the mass extinction of megafauna in the late Pleistocene. In “The Geographical Distribution of Animals” (1876) he wrote, “We live in a zoologically impoverished world, from which all the hugest, and fiercest, and strangest forms have recently disappeared”. He added that he believed the most likely cause for the rapid extinctions to have been glaciation, but by the time he wrote “World of Life” he had come to believe those extinctions were “due to man’s agency”.In 1880, Wallace published the book “Island Life” as a sequel to “The Geographical Distribution of Animals.” It surveyed the distribution of both animal and plant species on islands. Wallace classified islands into three different types. Oceanic islands, such as the Galapagos and Hawaiian Islands; then known as the Sandwich Islands, formed in mid ocean region and never part of any large continent. Such islands were characterized by a complete lack of terrestrial mammals and amphibians, and their inhabitants; with the exceptions of migratory birds and species introduced by human activity, were typically the result of accidental colonization and subsequent evolution. He divided continental islands into two separate classes depending on whether they had recently been part of a continent; like Britain, or much less recently like Madagascar. He discussed how those 3 differences affected the flora and fauna. He detailed how isolation affected evolution and how that could result in the preservation of classes of animals, such as the lemurs of Madagascar that were remnants of once widespread continental faunas. He extensively discussed how changes of climate, particularly periods of increased glacial activity, may have affected the distribution of flora and fauna on some islands, and the first portion of the book discusses possible causes of these great ice ages. “Island Life” was considered a very important work at the time of its publication. It was discussed extensively in scientific circles both in published reviews and in private correspondence.
Environmental Issues:
Wallace’s extensive work in biogeography made him aware of the impact of human activities on the natural world. In “Tropical Nature” and Other Essays (1878), he warned about the dangers of deforestation and soil erosion, especially in tropical climates prone to heavy rainfall. He noted the complex interactions between vegetation and climate warning that the extensive clearing of rainforest for coffee cultivation in Ceylon (Sri Lanka) and India would adversely impact the climate in those countries and lead to their eventual impoverishment due to soil erosion. In “Island Life,” Wallace again mentioned deforestation and also the impact of invasive species. On the impact of European colonization on the island of Saint Helena, he wrote:
“… yet the general aspect of the island is now so barren and forbidding that some persons find it difficult to believe that it was once all green and fertile. The cause of this change is, however, very easily explained. The rich soil formed by decomposed volcanic rock and vegetable deposits could only be retained on the steep slopes so long as it was protected by the vegetation to which it in great part owed its origin. When this was destroyed, the heavy tropical rains soon washed away the soil and have left a vast expanse of bare rock or sterile clay. This irreparable destruction was caused, in the first place, by goats, which were introduced by the Portuguese in 1513, and increased so rapidly that in 1588 they existed in the thousands. These animals are the greatest of all foes to trees, because they eat off the young seedlings, and thus prevent the natural restoration of the forest. They were, however, aided by the reckless waste of man. The East India Company took possession of the island in 1651, and about the year 1700 it began to be seen that the forests were fast diminishing, and required some protection. Two of the native trees, redwood and ebony, were good for tanning, and, to save trouble, the bark was wastefully stripped from the trunks only, the remainder being left to rot; while in 1709 a large quantity of the rapidly disappearing ebony was used to burn lime for building fortifications!”
Wallace’s comments on environment grew more strident later in his career. In 1911 in “The World of Life” he wrote:
“These considerations should lead us to look upon all the works of nature, animate or inanimate, as invested with certain sanctity, to be used by us but not abused, and never to be recklessly destroyed or defaced. To pollute a spring or a river, (and these days the air) to exterminate a bird or beast, should be treated as moral offences and as social crimes;…… Yet during the past century, which has seen those great advances in the knowledge of Nature of which we are so proud, there has been no corresponding development of a love or reverence for her works; so that never before has there been such widespread ravage of the earth’s surface by destruction of native vegetation and with it of much animal life, and such wholesale defacement of the earth by mineral workings and by pouring into our streams and rivers (air) the refuse of manufactories and of cities; and this has been done by all the greatest nations claiming the first place for civilization and religion!

It is little wonder that Wallace was awarded the medal from the Linnean society. His acute sense coupled with affection and devoutness of nature was displayed throughout his life with the wrong doings which he cites constantly are being perpetrated by us humans on nature manifests itself upon him that he is prepared to forego everything for their benefit.
Charles Darwin made 6 major statements on his theories about Evolution.
Over production or plagues occur in nature.
Not all the individual organisms can survive.
Variations occurs in every population.
Variations are inherited by the next generation.
Organisims with favourable variations will survive and reproduce accordingly.
Populations as a whole become fitter and evolve on these favourable variations.
The scientific inquiry into Darwin’s “the origin of species” can be dated to at least the 6th century BCE, with the Greek philosopher Anaximander (500 BC). He influenced Theophrastus in his works of nature, geography and Apeiron is spatially infinite. Only fragments of his work still exist despite being well documented at the time. Others who had considered evolutionary ideas included the Greek philosopher Empedocles, the Roman philosopher and poet Lucretius, the Afro-Arab biologist Al Jahiz, the Persian philosopher Ibn Miskawayh, the Brethren of Purity, and the Chinese philosopher Zhuangzi (Zhuang Zhou 400 BC. he was He did not pursue this line as pressure implied his basic theories on evolution were counterproductive to modern civilisation back then.). As biological knowledge grew the ideas of the biologist Jean-Baptiste Lamarck about transmutation of species influenced radicals, but were rejected by mainstream scientists.
Debate about the mechanisms of evolution continued. Darwin could not explain the source of the heritable variations which would be acted on by natural selection. Like Lamarck, he thought that parents passed on adaptations acquired during their lifetimes. In the 1880’s August Wesmann’s experiments indicated that changes from use to disuse were not heritable and Lamarck’s theory gradually fell out of favour.
More significantly, Darwin could not account for how traits were passed down from generation to generation. In 1865 Gregor Mendel discovered that traits were inherited in a very predictable manner. When Mendel’s work was rediscovered in the early 1900s, disputes over the rate of evolution predicted by early geneticists and biometricians led to a rift between the Mendel’s supporters and Darwin’s model on evolution.
Yet it was the rediscovery of Gregor Mendel’s pioneering work on the fundamentals of genetics; of which Darwin and Wallace were unaware; by Hugo de Vries and others in the early 1900s that provided the impetus for a better understanding of how variation occurs in both plant and animal traits. That variation is the main fuel used by natural selection to shape the wide variety of adaptive traits observed in organic life. Hugo de Vries and other early geneticists rejected gradual natural selection. The rediscovery of Gregor Mendel’s papers and experiments on genetics eventually provided a solid base and a mountain of facts on which the theory of evolution stood even more convincingly than when it was originally proposed.
The apparent contradiction between Darwin’s theory of evolution by natural selection and Mendel’s work was reconciled in the 1920s and 1930s by evolutionary biologists such as J.B.S. Haldane, Sewall Wright, and particularly Ronald Fisher, who set the foundations for the establishment of the field of population genetics. The end result was a combination of evolution by natural selection and Mendel’s theory of inheritance.
In the 1940s, Oswald Avery’s discovery and identification of DNA as the genetic material and the subsequent publication of the structure of DNA by James Watson and Francis Crick in 1953, demonstrated the physical basis for inheritance. Since then, genetics and molecular biology have become core parts of evolutionary biology and have revolutionized the field of phylogenetics.
Darwin presents his case:
Natural variation needed to be proved. Stating that life is the survival of the fittest and descent modifications would not suffice. Fossil records, geographical distribution of living organisms, homologous body structures coupled with the similarities of early foetal development were used to support the theories.
Darwin submitted an enormous quantity of evidence supporting every aspect that life has changed or evolved over a long time. He used Wallace’s theories and proposed hypothesis to explain how and why evolution occurs.
Natural variation and artificial selection:
This can be seen as differences in individual species within an organism.
* In nature inheritable differences or variation are common factors.
* During the period of Darwin’s explanation no one knew why variation occurred or how inheritance worked.
* But he could cite work done to produce more milk or more productive grains, larger fruits or even different coloured and size dogs. All these organisms show variable traits.
* Artificial Selection was being carried out by humans as he cited with dairy cows.
Cows that produced higher yields were bred together to producer better milk.
Darwin maintained that evolution by natural selection existed on a daily basis with daily struggles. He realized that high birth rates and shortage of life’s basic needs forced organisms into a constant struggle to survive. Only the fastest predators were able to catch their prey while the slowest starved. The same could be said for the prey the fastest escaped while the slowest were caught and eaten.
Not only was speed essential but fitness also ensured survival. An individual’s fitness ensured that they would survive in their specific niche and reproduce.
The dire need to adapt to changing situations ensured that the offspring would have the best chance to survive if their characteristics both physical and behavioural were those that adapted best to the changing circumstances.
High Fitness meant better survival techniques with more offspring.
Low Fitness meant poor survival techniques with fewer offspring.
Natural selection explains how over a period of time organisms become better suited to their environment as they respond to their selection of changing environmental pressures.
The Descent of Modification:
* For around 500 million years, natural selection has resulted in the inherited characteristics of a population that best suits its environment and continues today. These changes increased the fitness of populations to their new environment.
* All living organisms were produced gradually over long periods conditioning themselves to their environmental niches through a process of evolutionary change.
* Each organism is the result of its fore parents and other species ability to change and adapt to new conditions over a period of time.
* All living organisms have a common ancestor. The author’s views are contradictory to a single ancestor. My hypothesis is that there were several initial ancestors that developed probably over several thousand or even million years. These new life forms were closely associated but different so that they were able to share properties and this lead to the diversity and ability for adaptations in future generations.
The evidence was needed and found:
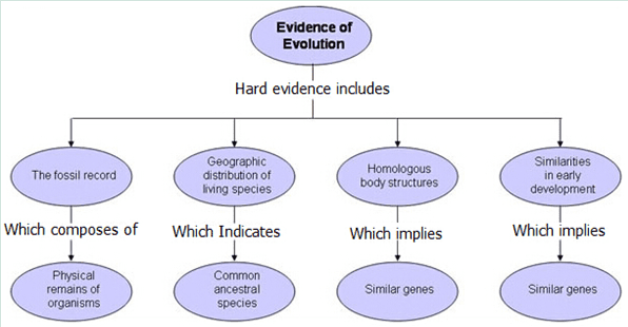
The evidence of evolution of Evolution was grouped into several categories including.
* Structural Adaptation: Developed over long periods, sometimes millions of years and include body parts, mimicry and Camouflage.
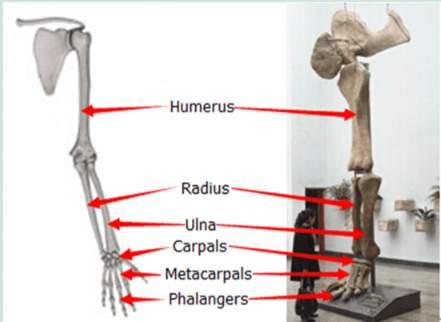
Each limb has adapted in ways that enabled the organisms to survive in different environments. These structural differences develop, as a result of the organism’s different use of that structure. Here the fore limb bones of birds, bats, whales, tortoises, crocodiles and kangaroos developed differently as a result of environmental conditions and different uses. However the limb bones all developed from the same clumps of cells in the growing embryonic stage.
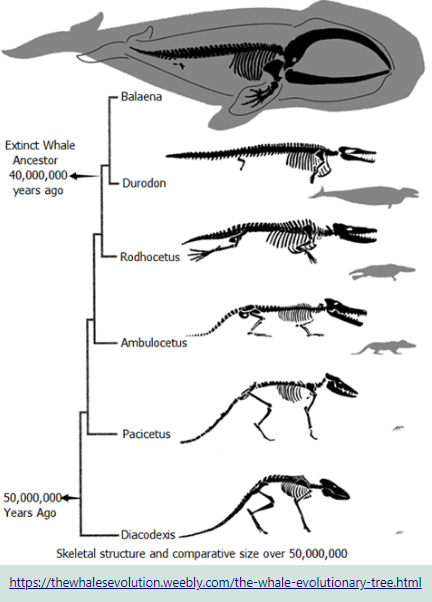
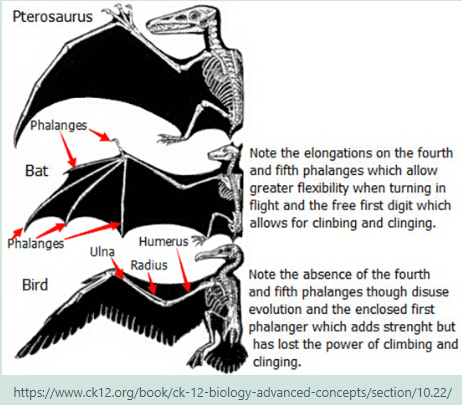
Homologous Structures are those structures that have the same embryonic tissues but develop different forms as they mature. The similarities and differences in bone structures, placement and functions assist the biologist in determining the organisms class according to how recently they last shared a common ancestor.
Anologous Structures are those structures that have different evolutionary embryonic tissues and remain different as they mature.
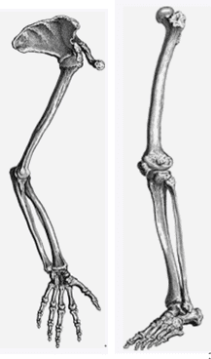
Above: Amazing, even after 65 to 70 million years animals still have the same basic bone structures as can be seen from these Dinosaur bones on display adjacent to the Zi Gong Dinosaur fossil pits in south western Sichuan China.
Below: Human arm bone structure left and leg bone structure right still have the same components just differing in size.
Structural adaptations include mimicry and camouflage. Mimicry is where one animal superficially resembles another in appearance, scent or colour but is not related. Many native orchids and lilies mimic the scents of female wasps in order to attract male wasps in order to carry out pollination.
Camouflage is where an organism has the traits of blending into its surroundings while the opposite is also true where the adapted traits are very conspicuous and acts as a warning sign to others.

Photographed in The Orchis gardens of Charles Darwin England. These Digger Wasps are attracted to the flower by powerful pheromone scents the orchids give off to attract pollinators in which the male wasps kindly but confusingly oblige.
* Adaptive Radiation: occurs over medium time periods on the evolution scale of thousands of years or a few million years. The species evolve from another species to fit into various habitat niches. The honeyeaters of Australia have just a few common ancestors yet the individual species now number over a hundred. The Asian dog is the common ancestor of the Australian Dingo, which was bought to Australia or migrated to Australia within the last 50,000 years ago. (some research now indicates that the Dingo may have been in Australia for as little as 5,000 years.) As the continent again became isolated after the last Ice Age, conditions became drier and harsher. This resulted in the smaller, leaner faster dog compared to the wolves with greater stamina, which was acquired to survive while adapting better in order to exist in the new environment.

* Physiological Adaptation: These adaptations develop over relatively short periods, within a few generations and include antibiotic resistance by bacteria.
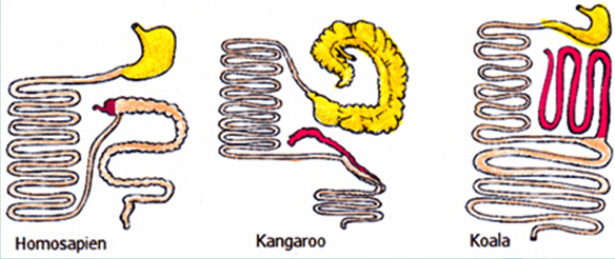
A Vestigial Structure has no present useful function or has declined to a point that it is almost dysfunctional and not necessary to the species. The human appendix is now dysfunctional and small, the appendix in the Koala is strongly developed.
* Fossil Records: Fossil records are useful in determining the chronological age of an organism’s existence, its habitat, structure and evolutionary process on how the species evolved to adapt to its new environment.
Fossils also depict climate and habits of organisms which can be compared to organisms that are still living today.
* Geographical Distribution: Species on the various Continents had all descended from common ancestors. As a result some species evolved similarly due to living in similar habitats with similar pressures. Since the breakup of Gondwanna the South American and Australian Mammals adapted differently but those where the habitats changed in similar ways the plants and animals also developed along parallel lines with similar traits. Here the Opossum and the Australian Brush Tail Possum exhibit similar traits. The example below shows strong hind leg muscles, small front legs good at clasping thick fur, large eyes for night vision and similar facial structures for feeding on fruits and leaves.

* Anatomy: Is the scientific study of the physical structure of animals. The position, shape and size of the different structures not only told biologists what an animal looked like but is essential in allowing biologists in determine how a specie moved, how fast or slow they moved, hunted or grazed its habitat niche and reproduction methods.
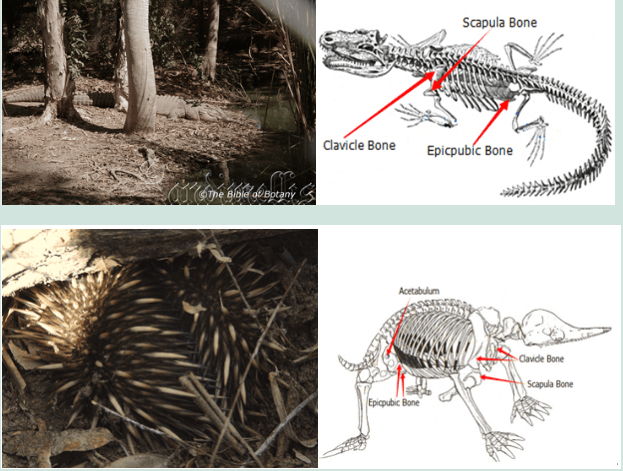
Echidnas the darlings of the Australian bush are a link between reptiles and marsupial mammals.
Their bone structures have many similarities. The Clavical, scapula and Epicpubic bone structures are very similar in monotremes and reptiles which explains the way they walk, more like a swimming walk found in crocodiles and goannas.
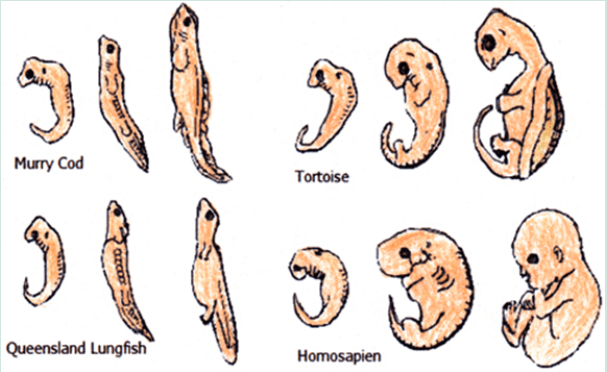
* Embryology: The early stages of most animals with backbones are so alike that they defy the average person any differences to tell them apart. The same group of embryonic cells will develop in the same manner to those produced the tissues, organs and bones of all vertebrates.
The earliest stages of all vertebrae organisms look very similar. All fish, amphibians, birds, mammals and marsupials have gills and tails as embryos.
* Biochemical: Is the latest methods for detailing organisms using chemical analysis and DNA. Detailed below.
To summarize the theories:
* Individual organisms in nature differ from one another. Some of this variation is inherited.
* All organisms in nature produce more offspring than can survive and many of those that do survive do not reproduce.
* If more organisms are produced than can virtually survive then members of that specie must compete for the limited resources available to it.
* If each organism is unique then each one has different advantages and possible disadvantages to aid its struggle for existence and survival of the species.
* The individual which are best adapted to their environmental niche, survive and reproduce to further add to the fitness of their species in the struggle to survive. Their characteristics are passed onto their offspring. Those individuals that are not well adapted to their environment will not survive or leave fewer offspring further diminishing the opportunities for survival.
* Species change over long periods of time due to natural selection of the fittest.
* Species that are living today are the results of modifications made from species that lived in the distant past.
* All the organisms on Earth today are united into a single tree of which there is a common ancestor.
How does Modern Science fit into Darwin
and Wallace’s Theories on Evolution:
The following article is rewritten by the author for simplicity form an article taken from nhm.ac.uk/nature-online/evolution,
In its early history, evolutionary biology primarily drew in scientists from traditional taxonomically oriented disciplines, whose specialist training in particular organisms addressed general questions in evolution. Evolutionary biology expanded as an academic discipline, particularly after the development of the modern evolutionary synthesis. It began to draw more widely from the biological sciences. Currently the study of evolutionary biology involves scientists from fields as diverse as biochemistry, ecology, genetics and physiology combined with evolutionary concepts. They are used in even more distant disciplines such as psychology, medicine, philosophy and computer science. In the 21st century, current research in evolutionary biology deals with several areas where the modern evolutionary synthesis may need modification or extension, such as assessing the relative importance of various ideas on the unit of selection and evolvability and how to fully incorporate the findings of evolutionary developmental biology.
Heredity, DNA & Evolution:
Evolution in organisms occurs through changes in traits that are passed down from the parents in particular characteristics of an organism to help it adapt to its environment. In humans the eye colour is an inherited characteristic passed down from one of their parents. This will result in brown, blue or green eyes. Inherited traits are controlled by genes and the complete set of genes within an organism’s genome is called its genotype while the complete set of observable traits that make up the structure and behavior of an organism is called its phenotype.
These traits come from the interaction of its genotype with the environment. As a result, many aspects of an organism’s phenotype are not inherited. For example, suntanned skin comes from the interaction between a person’s genotype and sunlight; thus, suntans are not passed on to people’s children as is short scalpel hair. However, some people tan more easily than others, due to differences in their genotype; a striking example are people with the inherited trait of albinism, who do not tan at all and are very sensitive to sunburn.
Heritable traits are passed from one generation to the next via DNA, a molecule that encodes genetic information. The DNA is a long polymer composed of four types of bases. The sequence of the bases is a particular DNA molecule with specific genetic information. It is similar to a sequence of letters spelling out words and a sentence. DNA is heritable because the specific pairing of the four bases provides a biochemical mechanism that cells use to accurately transcribe and replicate the coded information from one parent to its offspring.
Portions of a DNA molecule that specify a single functional unit are called genes; different genes have different sequences of bases. Within cells, the long strands of DNA form condensed structures called chromosomes. A specific location within a chromosome is known as a locus. If the DNA sequence at a locus varies between individuals, the different forms of this sequence are called alleles.
DNA sequences can change through mutations, producing new alleles. If a mutation occurs within a gene, the new allele may affect the trait that the gene controls, altering the phenotype of the organism.
However, while this simple correspondence between an allele and a trait works in some cases, most traits are more complex and are controlled by multiple interacting genes.
The study of such complex traits is a major area of current genetic research. Another unsolved question in genetics is epigenetics. This is important in evolution as this is where heritable changes occur in organisms without there being any changes to the sequences of their genes.
Variation:
An individual organism’s phenotype results from both its genotype and the influence from the environment it has lived in. A substantial part of the variation in phenotypes in a population is caused by the differences between their genotypes. The modern evolutionary synthesis defines evolution as change over time in this genetic variation. The frequency of one particular allele will fluctuate, becoming more or less prevalent relative to other forms of that gene. Evolutionary forces act by driving these changes in allele frequency in one direction or another. Variation disappears when a new allele reaches the point of fixation. That is when it either disappears from the population or replaces the ancestral allele entirely.
Variation comes from mutations in genetic material, migration between populations (gene flow), and the reshuffling of genes through sexual reproduction. Variation also comes from exchanges of genes between different species; for example, through horizontal gene transfer in bacteria, and hybridization in plants.[39] Despite the constant introduction of variation through these processes, most of the genome of a species is identical in all individuals of that species. However, even relatively small changes in genotype can lead to dramatic changes in phenotype: chimpanzees and humans differ in only about 5mm of their genomes.
Mutation:
Random mutations constantly occur in the genomes of organisms; these mutations create genetic variation. Mutations are changes in the DNA sequence of a cell’s genome and are caused radiation, viruses, transposons and mutagenic chemicals, as well as errors that occur during meiosis or DNA replication.
Mutations involve several different types of change in DNA sequences. They can have no effect, alter the product of a gene or prevent the gene from functioning.
Studies in the fly Drosophila melanogaster suggest that if a mutation changes a protein produced by a gene, this will probably be harmful, with about 70 percent of these mutations having damaging effects, and the remainder being either neutral or weakly beneficial. Due to the damaging effects that mutations can have on cells, organisms have evolved mechanisms such as DNA repair to remove mutations. Therefore, the optimal mutation rate for a species is a trade-off between costs of a high mutation rate, such as deleterious mutations, and the metabolic costs of maintaining systems to reduce the mutation rate, such as DNA repair enzymes. Viruses that use RNA as their genetic material have rapid mutation rates, which can be an advantage since these viruses will evolve constantly and rapidly and thus evade any defensive responses. That is they can evade natural or synthetic animal immune systems.
Mutations can involve large sections of a chromosome becoming duplicated; usually by genetic recombination, which can introduce extra copies of a gene into a genome. Extra copies of genes are a major source of the raw material needed for new genes to evolve.
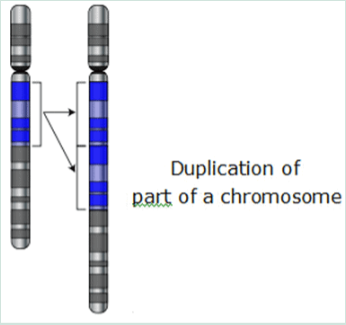
This is important because most new genes evolve within gene families. Preexisting genes that share common ancestors can be found in the human eye. The eyes have four genes to make structures that sense light. Three are for color vision and one for night vision yet all four are descended from a single ancestral gene.
New genes can be created from an ancestral gene when a duplicate copy mutates and acquires a new function. This process is easier once a gene has been duplicated because this increases redundancy; with one gene in the pair acquiring a new function while the other copy still performs its original function.
Other types of mutation can even create entirely new genes from previously non coding DNA. The creation of new genes can also involve small parts of several genes being duplicated, with these fragments then recombining to form new combinations with new functions.
When new genes are assembled from shuffling pre-existing parts, domains act as modules with simple independent functions, which then can be mixed together creating new combinations with new and complex functions. Polyketide synthases are large enzymes that make antibiotics and contain up to one hundred independent domains that each catalyze one step in the overall process. Look at it like a single stage in a processing plant.
Changes in chromosome number may involve even larger mutations, where segments of the DNA within chromosomes break and then rearrange. For example, two chromosomes in the Homo genus fused to produce human chromosome 2; this fusion did not occur in the lineage of the other apes, and they retain these separate chromosomes. In evolution, the most important role of such chromosomal rearrangements may be to accelerate the divergence of a population into new species by making populations less likely to interbreed, and thereby preserving genetic differences between these populations.
Sequences of DNA that can move about the genome, such as transposons, make up a major fraction of the genetic material of plants and animals, and may have been important in the evolution of genomes.[63] For example, more than a million copies of the Alu sequence are present in the human genome, and these sequences have now been recruited to perform functions such as regulating gene expression.[64] Another effect of these mobile DNA sequences is that when they move within a genome, they can mutate or delete existing genes and thereby produce genetic diversity.
Sex and recombination:
In asexual organisms, genes are inherited together, or linked, as they cannot mix with genes of other organisms during reproduction. In contrast, the offspring of sexually reproduced organisms contain random mixtures of their parents’ chromosomes that are produced through independent assortment.
In a related process called homologous recombination, sexual organisms exchange DNA between two matching chromosomes. Recombining and resorting do not alter the allele frequencies, but instead change which alleles are associated with each other, producing offspring with new combinations of alleles. Sex or pollination is a good way to increase genetic variation and may increase the rate of evolution.
However, asexuality is advantageous in some environments as it can evolve in previously-sexual animals. Here, asexuality might allow the two sets of alleles in their genome to diverge and gain different functions. Recombination allows even alleles that are close together in a strand of DNA to be inherited independently. However, the rate of recombination is low at approximately two events per chromosome per generation. As a result, genes close together on a chromosome may not always be shuffled away from each other, and genes that are close together tend to be inherited together, a phenomenon known as linkage. This tendency is measured by finding how often two alleles occur together on a single chromosome, which is called their linkage disequilibrium. A set of alleles that is usually inherited in a group is called a haplotype. This can be important when one allele in a particular haplotype is strongly beneficial. Natural selection can drive a selective sweep that will also cause the other alleles in the haplotype to become more common in the population. This effect is called genetic hitchhiking.
When alleles cannot be separated by recombination – such as in mammalian Y chromosomes, which pass intact from fathers to sons – harmful mutations accumulate. By breaking up allele combinations, sexual reproduction allows the removal of harmful mutations and the retention of beneficial mutations. In addition, recombination and reassortment can produce individuals with new and advantageous gene combinations. These positive effects are balanced by the fact that sex reduces an organism’s reproductive rate, can cause mutations and may separate beneficial combinations of genes. The reasons for the evolution of sexual reproduction are unclear and this question is still an active area of research in evolutionary biology.
Population Genetics:
From a genetic viewpoint, evolution is a “generation-to-generation change in the frequencies of alleles within a population that shares a common gene pool.” A population is a localized group of individuals belonging to the same species. For example, all of the moths of the same species living in an isolated forest represent a population. A single gene in this population may have several alternate forms, which account for variations between the phenotypes of the organisms. A gene for coloration in moths that has two alleles: black and white is found in the Pepper Moth. A gene pool is the complete set of alleles for a gene in a single population. The allele frequency measures what the fraction of the gene pool is composed of a single allele. That is what fraction of moth coloration genes are the black allele).
Evolution occurs when there are changes in the frequencies of alleles within a population of interbreeding organisms. The allele for black color in the Pepper Moth populations became more common.

To understand the mechanisms that cause a population to evolve, it is useful to consider what conditions are required for a population not to evolve. The “Hardy-Weinberg principle” states that the frequencies of alleles in a sufficiently large population will remain constant if the only forces acting on that population are the random reshuffling of alleles during the formation of the sperm or egg, and the random combination of the alleles in these sex cells during fertilization. Such a population is said to be in “Hardy-Weinberg equilibrium,” and is not evolving.
Gene Flow:
Gene flow is the exchange of genes between populations, which are usually of the same species. Gene flow within a species includes the migration and then sexual breeding of organisms or the exchange of pollen during pollination. Gene transfer between species includes the formation of hybrid organisms and horizontal gene transfer.
Migration into or out of a population can change allele frequencies, as well as introducing genetic variation into a population. Immigration may add new genetic material to the established gene pool of a population or conversely, emigration may remove genetic material.

in his territory. overly nutritious and the need to save energy.
When male koalas mature they will be forced to leave their mother and population by the resident bull. They will take over or muscle in on a new territory a new population. At first the territory will be small but as the bull gets stronger and larger the area will also become larger and more females will be acquired to mate with causing gene flow between populations. Weak bulls will have nowhere to go, no females to pass on the stronger more adaptable genes.
As barriers to reproduction between two diverging populations are required for the populations to become new species, gene flow may slow this process by spreading genetic differences between the populations. Gene flow is hindered by mountain ranges, oceans and deserts or even man-made structures such as the Great Wall of China, and the Suez Canal which has hindered the flow of plant genes.
Depending on how far two species have diverged since their most recent common ancestor, it may still be possible for them to produce offspring. Grevillea banksii and Grevillea bipinatifolia can be cross pollinated and produce offspring known as mules. Early produced mules were that of a horse and a donkey. Such hybrids are generally infertile, due to the two different sets of chromosomes being unable to pair up during meiosis.
In some cases more closely related species may regularly interbreed, but hybrids will be selected against and the species will remain distinct. However, viable hybrids are occasionally formed and these new species can either have properties intermediate between their parent species, or possess a totally new phenotype as is the case of Grevillea banksii and Grevillea pteridifolia. The importance of hybridization in creating new species of animals is unclear, although cases have been seen in many types of plants and animals. The hybrids usually are not well adapted to their environment and soon become extinct or are reabsorbed into one or the other parents.
Hybridization is, however, an important means of speciation in plants, since polyploidism (Is where a plant or animal has more than two copies of each chromosome) is tolerated in plants more readily than in animals. Polyploidy is important in hybrids as it allows reproduction, with the two different sets of chromosomes each being able to pair with an identical partner during meiosis. Polyploids also have more genetic diversity, which allows them to avoid in breeding depression in small populations.
Horizontal gene transfer is the transfer of genetic material from one organism to another organism that is not its offspring; this is most common among bacteria. In medicine, this contributes to the spread of antibiotic resistance, as when one bacteria acquires resistance genes from its host, it can rapidly transfer them to other species. Horizontal transfer of genes from bacteria to eukaryotes such as the yeast Saccharomyces cerevisiae and the adzuki bean beetle Callosobruchus chinensis may also have already occurred. Large scale transfers are known amongst the eukaryotic bdelloid rotifers, which appear to have received a range of genes from bacteria, fungi, and plants. Viruses can also carry DNA between organisms, allowing transfer of genes even across biological domains. Large-scale gene transfer has also occurred between the ancestors of eukaryotic cells and prokaryotes, during the acquisition of chloroplasts and mitochondria, of chloroplasts and mitochondria.
Mechanisms:
The two main mechanisms that produce evolution are natural selection and genetic drift. Natural selection favors genes that aid survival and reproduction. Genetic drift is random change in the frequency of alleles, caused by the random sampling of a generation’s genes during reproduction. The relative importance of natural selection and genetic drift in a population varies depending on the strength of the selection and the effective population size, which is the number of individuals capable of breeding. Natural selection usually predominates in large populations, while genetic drift dominates in small populations. The dominance of genetic drift in small populations can even lead to the fixation of slightly deleterious mutations. As a result, changing population size can dramatically influence the course of evolution. Population bottlenecks, where the population shrinks temporarily and therefore loses genetic variation, result in a more uniform population. It is also possible that overpopulation can create a circumstance where the population adapts to marginal habitats which were not occupied prior to the explusion.
Natural Selection:
Natural selection is the process by which genetic mutations that enhance reproduction become, and remain, more common in successive generations of a population. It has often been called a “self-evident” mechanism because it necessarily follows from three simple facts:
* Heritable variation exists within populations of organisms.
* Organisms produce more offspring than can survive.
* These offspring vary in their ability to survive and reproduce.
These conditions produce competition between other organisms and the species for survival and reproduction. Organisms with traits that are more adaptable and advantages over their competitors pass these traits on. Conversely organisms that do not confer an advantage are not passed on to the next generation and die out.
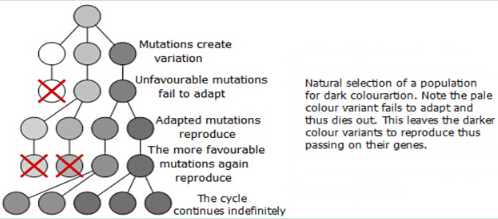
The central concept of natural selection is the evolutionary fitness of an organism. Fitness is measured by an organism’s ability to survive and reproduce, which determines the size of its genetic contribution to the next generation. Fitness is not the same as the total number of offspring but is indicated by the proportion of subsequent generations that carry an organism’s genes. If an organism could survive well and reproduce rapidly, but its offspring were all too small and weak to survive a drought or other natural phenomenon, this organism would make little genetic contribution to future generations and would have low fitness.
If an allele increases fitness more than the other alleles of that gene, then with each generation this allele will become more common within the population. These traits are said to be “selected for”. Examples of traits that can increase fitness are enhanced survival, and increased fecundity. Conversely the lower fitness, caused by having a less beneficial or deleterious allele may result in this allele becoming rarer and they are “selected against”. It is important that the fitness of an allele is not a fixed characteristic for if the environment changes, previously neutral or harmful traits may become beneficial and previously beneficial traits may become disadvantageous. Even if the direction of selection does reverse in this way, traits that were lost in the past may not re-evolve in an identical form.
Natural selection within a population for a trait can vary across a range of values, such as height. This can be categorized into three different types.
* The first is directional selection, which is a shift in the average value of a trait over time. An organism slowly grows taller.
* Secondly, disruptive selection is selection for extreme trait values and often results in two different values becoming most common, with selection against the average value. This would be when either the short or the tall organisms had an advantage, but not those of medium height.
* Finally, in stabilizing selection there is bias against the extreme trait’s values on both ends, which causes a decrease in variance around the average value and less diversity in the gene pool. This would cause organisms to slowly become all the same height.
A special case of natural selection is sexual selection, which is selection for any trait that increases mating success by increasing the attractiveness of an organism to potential mates usually the female. Traits that evolved through sexual selection are particularly prominent in males of some animal species, despite traits such as cumbersome antlers and claws, mating calls or bright colors that attract predators and consequently decrease the survival rates of individual males. This survival disadvantage is balanced by higher reproductive success in males that show these hard to fake, sexually selected traits.
Natural selection most generally makes nature the measure against which individuals, and individual traits, are more or less likely to survive. “Nature” in this sense refers to an ecosystem where organisms interact with every other element, physical as well as biological, in their local environment. Eugene Odum, a founder of ecology, defined an ecosystem as: “Any unit that includes all of the organisms…in a given area interacting with the physical environment so that a flow of energy leads to a clearly defined trophic structure, biotic diversity, and material cycles (That is the exchange of materials between living and nonliving segments) within the system.” Each population within an ecosystem occupies a distinct niche, or position, with distinct relationships to other parts of the system. These relationships involve the life history of the organism, its position in the food chain, and its geographic range. This broad understanding of nature enables scientists to delineate specific forces which, together, comprise natural selection.
An active area of research is the unit of selection, with natural selection being proposed to work at the level of genes, cells, individual organisms, groups of organisms and species. None of these are mutually exclusive and selection can act on multiple levels simultaneously. When selection occurs below the level of the individual organism genes called transposons, which can replicate and spread throughout a genome. Selection at a level above the individual, such as group selection, may allow the evolution of co-operation.
Genetic Drift:
Genetic drift is the change in allele frequency from one generation to the next that occurs because alleles in offspring is a random sample of those in the parents, as well as from the role that chance plays in determining whether a given individual will survive and reproduce. In mathematical terms, alleles are subject to sampling error. As a result, when selective forces are absent or relatively weak, allele frequencies tend to “drift” upward or downward randomly in what is known as a random walk. This drift halts when an allele eventually becomes fixed either by disappearing from the population, or replacing the other alleles entirely. Genetic drift may therefore eliminate some alleles from a population due to chance alone. Even in the absence of selective forces, genetic drift can cause two separate populations that began with the same genetic structure to drift apart into two divergent populations with different sets of alleles.
The time for an allele to become fixed by genetic drift is to a degree dependant on population size. Fixation occurs more rapidly in smaller populations than in larger populations. The precise measure of population that is important is called the effective population size. The effective population is always smaller than the total population since it takes into account factors such as the level of inbreeding, the number of animals that are too old or young to breed, and the lower probability of animals that live far apart managing to mate with each other.
Genetic drift can be seen in the loss of pigments within animals that live in caves or underground, a change that produces no obvious advantage or disadvantage to the organism in complete darkness. However, it is usually difficult to measure the relative importance of selection and drift, so the comparative importance of these two forces in driving evolutionary change is an area of current research. These investigations were prompted by the neutral theory of molecular evolution, which proposed that most evolutionary changes are the result of the fixation of neutral mutations that do not have any immediate effects on the fitness of an organism.
Most genetic changes occur in a population that are the result of constant mutation pressure and genetic drift. This form of the neutral theory is now largely abandoned, since it does not seem to fit the genetic variation seen in nature. However, a more recent and better-supported version of this model is the nearly neutral theory, where most mutations only have small effects on fitness.
Divergent Evolution: The term divergentis from the Latin divergere, meaning “go in different directions”. Evolution, in turn, came from Latin evolutionem, from evolvere, meaning “to unroll”. The evolution of a species is largely influenced by environmental factors and predation activities. It is the process, in which a single species diverged into two or more varieties then subspecies and eventually into new species. The species environments, habitats or niches may vary, which in return the individuals with the most suitable characteristics to suit the new variable conditions will be accentuated, displacing the existing characteristics. Divergent evolution may also pertain to the process of tracing back two or more species to their common ancestor and knowing how and why these separate species evolved or diverged. convergent and parallel.
Below Boronia rosmarinifolia displaying genetic variation in flower colour and leaf width could eventually lead to sub species followed by a new species given time and environmental changes or differences like soil or climate change.


Convergent Evolution: Convergent evolution (Con/com , which are Latin and Ancient Greek for coming together) is influenced when two distinctly different species or genre converge to take on similar visual characteristics because they are growing in the same type of environment, habitat, niche and may have the same predators.


Outcomes:
Evolution influences every aspect of the form and behavior of organisms. Most prominent are the specific behavioral and physical adaptations that are the outcome of natural selection. These adaptations increase fitness by aiding activities such as finding food, avoiding predators or attracting mates. Organisms can also respond to selection by co-operating with each other, usually by aiding their relatives or engaging in mutually beneficial symbiosis. In the longer term, evolution produces new species through splitting ancestral populations of organisms into new groups that cannot or will not interbreed.
These outcomes of evolution are sometimes divided into macro evolution which is evolution that occurs at or above the level of species, such as extinction and speciation, and micro evolution, which is smaller evolutionary changes, such as adaptations, within a species or population.
In general, macroevolution is regarded as the outcome of long periods of microevolution. Thus, the distinction between micro evolution and macro evolution is not a fundamental one. The difference is simply the time period involved.
However, in macroevolution, the traits of the entire species may be important. For instance, a large amount of variation among individuals allows a species to rapidly adapt to new habitats, lessening the chance of it going extinct, while a wide geographic range increases the chance of speciation, by making it more likely that part of the population will become isolated. In this sense, micro evolution and macro evolution might involve selection at different levels, with micro evolution acting on genes and organisms, versus macro evolutionary processes such as species selection acting on entire species and affecting their rates of speciation and extinction.
A common misconception is that evolution has goals or long-term plans. Evolution has no long term goal and does not necessarily produce greater complexity. Although complex species have evolved, this occurs as a result of the side effects of the overall number of organisms increasing with simple forms of life remaining more common.
The overwhelming majority of species are micro scopic prokaryotes, which form about half the world’s biomass despite their small size, and constitute the vast majority of the Earth’s biodiversity. Simple organisms have therefore been the dominant form of life on Earth throughout its history and continue to be the main form of life to the present day. Complex life only appears more diverse because it is more readily noticeable. The evolution of microorganisms is particularly important to modern evolutionary research, since their rapid reproduction allows the study of experimental evolution and the observation of evolution and adaptation in real time.
Adaptation:
Adaptation is one of the basic phenomena of biology and the basis of both Darwin and Wallace’s theories on evolution. It is the process whereby an organism becomes better suited to its habitat by adapting to it. The term adaptation may also refer to a trait that is important for an organism’s survival. The incessant grinding of grass by the teeth of herbivores or the ability of herbivores to run quickly to escape its predators is no accident.
* Adaptation is the evolutionary process whereby an organism becomes better able to live in its habitat or habitats.
* Adaptedness is the state of being adapted: the degree to which an organism is able to live and reproduce in a given set of habitats.
* An adaptive trait is an aspect of the developmental pattern of the organism which enables or enhances the probability of that organism surviving and reproducing.
Adaptation may cause either the gain of a new feature, or the loss of an ancestral feature. An example that shows both types of change is bacterial adaptation to antibiotic selection, with genetic changes causing antibiotic resistance by either modifying the target of the drug, or increasing the activity of transporters that pump the drug out of the cell. The bacteria Eschellaria coli has evolved the ability to use citric acid as a nutrient in a long a term laboratory experiment, Flavo bacterium has evolved a new enzyme that allows these bacteria to grow on the biproducts of nylon manufacturing and the soils bacterium Sphingbium sp. evolving an entirely new metabolic pathway that degrades the synthetic pesticide pentachlorophenol.
An interesting but still controversial idea is that some adaptations might increase the ability of organisms to generate genetic diversity and adapt by natural selection thus increasing an organisms chances to evolve more quickly.
So adaptation occurs through the gradual modification of existing structures. Consequently, structures with similar internal organization may have different functions in related organisms. This is the result of a single ancestral structure being adapted to function in different ways. The bones within a bat’s wings, for example, are very similar to those in mice feet and primate hands, due to the descent of all these structures from a common mammalian ancestor.
Since all living organisms are related to some extent, even organs that appear to have little or no structural similarity, such as the eyes in arthropod, squid and vertebrate or the limbs and wings of arthropods and vertebrates, can depend on a common set of homologous genes that control their assembly and function. This is called deep homology.
A Balenea whale skeleton, the flipper bones were adapted from the front leg bones while the vestigial leg bones of its ancestors are still present.
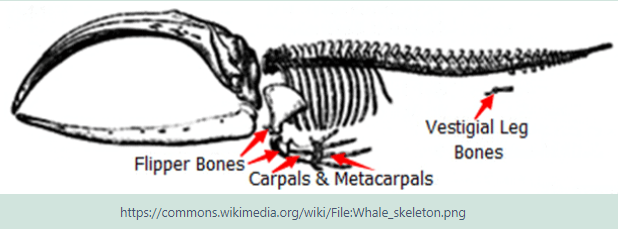
A Balenea whale skeleton, flipperbones, which were adapted from front leg bones, while remnant vestigial leg bones were the hind legs of its ancestors.
During adaptation, some structures may lose their original function and become “vestigial structures.” Such structures may have little or no function in a current species, yet had a clear function in its ancestral parents, or other closely related species. Examples include pseudogenes, the non-functional remains of eyes in blind cave dwelling fish, wings on flightless birds like the emu and the presence of hip bones in whales and snakes. Even, we humans are carrying around vestigial structures which include our wisdom teeth remnants of a more herbaceous diet, the coccyx bone the remnant of our once important tail, and the vermiform appendix which is more common and herbaceous animals that have a consumption of entirely green matter.
Many traits that appear to be simple adaptations are in fact exaptations. These structures were originally adapted for one function but coincidentally became somewhat useful for some other function in the process.

The British Flying Squirrel left and the Australian sugar Glider are good examples of how a flattened head and body allowed the pair to conceal themselves between trunks and bark, utilize small hollows but also assisted in the aid of gliding from tree to tree.
Within cells, molecular machines such as the bacterial flagella and protein sorting evolved by the recruitment of several pre existing proteins that previously had different functions. Another example is the recruitment of enzymes from glycolysis and xenobiotic metabolism to serve as structural proteins called crystallins within the lenses of organisms’ eyes.
A critical principle of ecology is that of competitive exclusion. Competitive expulsion dictates that no two species can occupy the same niche in the same environment for a long time. As a result of natural selection one of the species will succumb to the other or have to adapt to a different niche in the habitat. This means; for example, that the three species of wombats in Australia have all adapted to live in slightly different habitats, which minimizes the competition between them for food and shelter.
An area of current investigation in evolutionary developmental biology is the developmental basis of adaptations and exaptations. This research addresses the origin and evolution of embryonic development and how modifications of development and developmental processes produce novel features. Studies have shown that evolution can alter development to create new structures, such as embryonic bone structures that develop into a jaw in some animals while they form part of the middle ear in mammals.
It is also possible for structures that have been lost in evolution to reappear due to changes in developmental genes. This has been seen in the mutation of chickens causing embryos to grow teeth similar to those of ancestral reptiles similar to those of crocodiles. It is now becoming clear that most alterations in the form of organisms are due to changes in a small set of conserved genes.
Coevolution:
Interactions between organisms can produce both conflict and cooperation. When the interaction is between pairs of species, such as a pathogen and a host, or a predator and its prey, these species can develop matched sets of adaptations. Here, the evolution of one species causes adaptations in a second species. These changes in the second species then, in turn, cause new adaptations in the first species. This cycle of selection and response is called coevolution.
An example is the production of the advanced appendix of the koala to extract cyanide from gum leaves. In this herbivore plant pair, an evolutionary arms race has produced higher levels of cyanide to the disadvantage of all other warm blooded herbivores with correspondingly advancements in the koals ability to eliminate the toxins.
Cooperation:
However, not all interactions between species involve conflict. Many cases of mutually beneficial interactions have evolved. For instance, an extreme cooperation exists between plants and the mycorrhizal fungi that grow on their roots and aid the plant in absorbing nutrients from the soil. This is a reciprocal relationship as the plants provide the fungi with sugars from photosynthesis. Here, the fungi actually grow inside plant cells, allowing them to exchange nutrients with their hosts, while sending signals that suppress the plant immune system.
Coalitions between organisms of the same species have also evolved. An extreme case is the eusociality found in social insects, such as bees, termites and ants, where sterile insects feed and guard the small number of organisms in a colony that are able to reproduce.
On an even smaller scale, the somatic cells that make up the body of an animal limit their reproduction so they can maintain a stable organism, which then supports a small number of the animal’s germ cells to produce offspring. Here, somatic cells respond to specific signals that instruct them whether to grow, remain as they are, or die. If cells ignore these signals and multiply inappropriately, their uncontrolled growth causes cancer.
Such cooperation within species may have evolved through the process of kin selection, which is where one organism acts to help raise a relative’s offspring “kinship.” This activity is selected for because if the helping individual contains alleles which promote the helping activity, it is likely that its kin will also contain these alleles and thus those alleles will be passed on. Other processes that may promote cooperation include group selection, where cooperation provides benefits to a group of organisms.
Speciation:
Speciation is the process where a species diverges into two or more descendant species. The exact definition of the term “species” is still controversial, particularly in prokaryotes, and this is called the species problem. Biologists have proposed a range of more precise definitions, but the definition used is a pragmatic choice that depends on the particularities of the species concerned. Typically the actual focus on biological study is the population, an observable interacting group of organisms, rather than a species, an observable similar group of individuals.
Speciation has been observed multiple times under both controlled laboratory conditions and in nature. In sexually reproducing organisms, speciation results from reproductive isolation followed by genealogical divergence. There are four mechanisms for speciation.
The most common in animals is allopatric speciation, which occurs in populations initially isolated geographically, such as by habitat fragmentation or migration. Selection under these conditions can produce very rapid changes in the appearance and behaviour of organisms. As selection and drift act independently on populations isolated from the rest of their species, separation may eventually produce organisms that cannot interbreed.
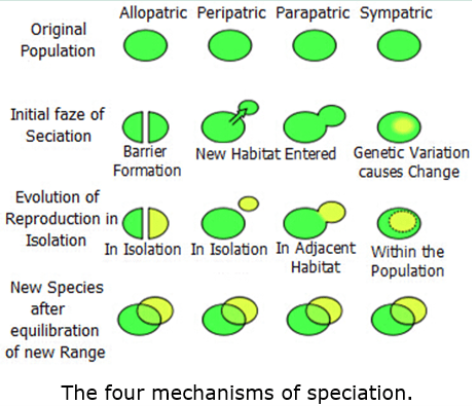
The second mechanism of speciation is peripatric speciation, which occurs when small populations of organisms become isolated in a new environment. This differs from allopatric speciation in that the isolated populations are numerically much smaller than the parental population. Here, the founder effect causes rapid speciation through both rapid genetic drift and selection on a small gene pool.
The third mechanism of speciation is parapatric speciation. This is similar to peripatric speciation in that a small population enters a new habitat, but differs in that there is no physical separation between these two populations. Instead, speciation results from the evolution of mechanisms that reduce gene flow between the two populations. Livistona marei and Livistona rigidula are a good example of parapatric speciation in process.
Generally this occurs when there has been a drastic change in the environment within the parental species’ habitat. One example is the grass Anthoxanthum odoratum, which can undergo parapatric speciation in response to localized metal pollution from mines. Here, plants evolve that have resistance to higher levels of metals in the soil. Selection against interbreeding with the metal-sensitive parental population produced a gradual change in the flowering time of the metal resistant plants, which eventually produced complete reproductive isolation. Selection against hybrids between the two populations may cause reinforcement, which is the evolution of traits that promote mating within a species, as well as character displacement, which is when two species become more distinct in appearance.

Geographical isolation of a finch on the Galapogos Islands produced over a dozen new species based on their food and possibly predator niches. The thicker, stronger bills would be better at adapting to larger, harder seeds, while the smaller more pointed beak would have adapted to smaller, softer seeds.
Finally, in sympatric speciation species diverge without geographic isolation or changes in habitat. This form is rare since even a small amount of gene flow may remove genetic differences between parts of a population. Generally, sympatric speciation in animals requires the evolution of both genetic differences and non random mating, to allow reproductive isolation to evolve.
One type of sympatric speciation involves cross breeding of two related species to produce a new hybrid species. This is not common in animals as animal hybrids are usually sterile. This is because during meiosis the homologous chromosomes from each parent are from different species and cannot successfully pair. However, it is more common in plants because plants often double their number of chromosomes, to form polyploids. This allows the chromosomes from each parent’s species to form a matching pair during meiosis, since as each parent’s chromosomes are represented by a pair already. An example of such a speciation event is when Arabidopsis atheliana and Arabidopsis arenosa cross-bred to give the new species Arabidopsis suecica. This happened about 20,000 years ago,[203] and the speciation process has been repeated in the laboratory, which allows the study of the genetic mechanisms involved in this process. Chromosome doubling within a species may be a common cause of reproductive isolation, as half the doubled chromosomes will be unmatched when breeding with undoubled organisms. Speciation events are important in the theory of punctuated equilibrium, which accounts for the pattern in the fossil record of short “bursts” of evolution interspersed with relatively long periods of stasis, where species remain relatively unchanged. In this theory, speciation and rapid evolution are linked, with natural selection and genetic drift acting most strongly on organisms undergoing speciation in novel habitats or small populations. As a result, the periods of stasis in the fossil record correspond to the parental population, and the organisms undergoing speciation and rapid evolution are found in small populations or geographically restricted habitats, and therefore rarely being preserved as fossils.
Extinction:
An organism is considered extinct when the entire species has disappeared or there is no visible proof of it still being in existence for a number of years. This includes any organism that may still be alive with the last remaining organism/s being unable to reproduce for a number of years since the last breeding pair was noted.
Extinction is not an unusual event, as species regularly appear through speciation, and disappear through extinction. Nearly all animal and plant species that have lived on earth are now extinct and extinction appears to be the ultimate fate of all species. These extinctions have happened continuously throughout the history of life. The rate of extinction peaks in occasional mass extinction events. The most well-known mass extinction was during the Cretaceous–Tertiary period that witnessed the mass extinction of all the dinosaurs except the future avian group. The earlier Permian–Triassic extinction event was even more severe, with approximately 96 percent of all species driven to extinction.


The Holocene extinction event is an ongoing mass extinction associated with humanity’s expansion across the globe over the past few thousand years with an expediential rate of extinctions being experienced at the present time. Present extinction rates are 100 to1000 times greater than the background rate, and up to 30 percent of species may be extinct by the mid 21st century. Human activities are now the primary cause of the ongoing extinction event; global warming will exacerbate the situation ensuring the present expediential growth will increase will continue well into the future.
The role of extinction in evolution is not very well understood and may depend on which type of extinction is considered. The causes of continuous “low-level” extinction events are still in its infancy as a science. These are the majority of extinctions and occur as an on-going event. They may be the result of competition between species for limited resources which we looked at as competitive exclusion. If one species can out compete another, this could produce species selection, with the species that is fitter surviving at the expense of the other. The intermittent mass extinctions are also important, but instead of acting as a selective force, they drastically reduce diversity in a nonspecific manner and promote bursts of rapid evolution and speciation in survivors.
Homosapiens rise is much like an extended time plague that will have catastrophic repercussions for the species and the environment at large. The small island pockets that do survive will then expand outwards from these islands and again re colonize the planet as per previous extinctions. To what role homosapien plays in the new earth will wholly depend on his ability to adapt or survive in isolated pockets probably as nomadic farmers. It is realistic to believe that this may happen as many other advanced beings like homo erectus, Peking man, Java Man man all became extinct for one reason or another.
Origin of life:
The origin of life is a necessary precursor for biological evolution, but understanding that evolution occurred once organisms appeared and investigating how this happens does not depend on understanding exactly how life began. The current scientific consensus is that the complex biochemistry that makes up life came from simpler chemical reactions, but it is unclear how this occurred. Not much is certain about the earliest developments in life, the structure of the first living things, or the identity and nature of any last universal common ancestor or ancestral gene pool. Consequently, there is no scientific consensus on how life began, but proposals include self replicating molecules such as RNA, and the assembly of simple cells.
Common descent:
All organisms on Earth are descended from a common ancestor or ancestral gene pool. The current species are in a flux of change in relation to their diversity which is a result of a long series of speciation and extinction events. The common descent of organisms was first deduced from four simple facts about organisms:
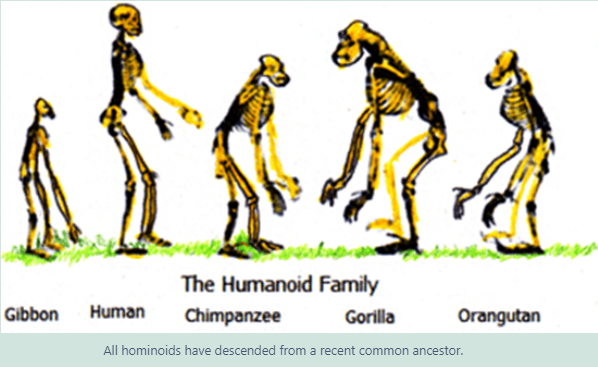
* They have geographic distributions that cannot be explained by local adaptation.
* The diversity of life is not a set of completely unique organisms, but organisms that share morphological similarities.
* Vestigial traits with no clear purpose resemble functional ancestratraits, and finally, that organisms can be classified using these similarities into a hierarchy of nested groups, similar to the construction of a family tree. Modern research suggests that, due to horizontal gene transfer the evolutionary tree is more complicated than the simple branching of a ancestral family tree. Some genes have spread independently between distantly related specie.
Past species have also left records of their evolutionary history. Fossils found around the world along with the “comparative anatomy” of present day organisms, constitute the morphological or anatomical, record.
Comparing the anatomies of present day organisms, chronologically recently extinct organisms to ancient extinctions species, paleontologists are able to infer the lineages of those species.
This approach is most successful for organisms that had hard body parts, such as shells, bones or teeth. It must be remembered with prokaryotes such as bacteria and archaea share a limited set of common morphologies, their fossils do not provide information on their ancestry.
Recently collated evidence for common descent has come from the study of biochemical similarities between organisms. For example, all living cells use the same basic set of nucleotides and amino acids. The development of molecular genetics has revealed the record of evolution left in organisms’ genomes. Dating species when they diverged through the molecular clock produced by mutations has confirmed direct relationships for common lineages. For example, these DNA sequence comparisons have revealed that humans and chimpanzees share 96mm of their genomes. By analyzing the few areas where they differ helps shed light on when the common ancestor of these species coexisted.
Evolution of life
Despite the uncertainty on how life began, it is generally accepted that prokaryotes inhabited the Earth from approximately 3.5 to 4 billion years ago. No obvious changes in morphology or cellular organization occurred in these organisms over the next few billion years.
The eukaryotes were the next major change in cell structure. These came from ancient bacteria being engulfed by the ancestors of eukaryotic cells, in a cooperative association known as endosymbiosis. The engulfed bacteria and the host cell then underwent co-evolution, with the bacteria evolving to form either mitochondria or hydrogenosomes. An independent second engulfment of cyanobacteria like organisms led to the formation of chloroplasts in algae and plants. (See article on The Science of plants chapter1.) It is unknown when the first eukaryotic cells appeared though they were well established between 1.6 billion and 2.7 billion years ago.
The history of life was that of the unicellular eukaryotes, prokaryotes, and archaea until about 610 million years ago. Somewhere around 610 million years ago multicellular organisms began to appear in the oceans in the Ediacaran period. The evolution of dependant multicellular organisms occurred in multiple, independent events, in organisms as diverse as sponges, brown algae, cyanobacteria, slime algae and myxobacteria.
Soon after the emergence of these first multicellular organisms, a remarkable amount of biological diversity appeared over approximately 10 million years, in an event called the Cambrian explosion. This is the period where the majority of modern day organisms appeared in the fossil records. These records showed many unique lineages that soon became extinct unable to adapt to the fast changing habitats. Various triggers for the Cambrian explosion have been proposed, including the accumulation of oxygen in the atmosphere due to the explosion of green producer plants utilizing photosynthesis.
About 500 million years ago, plants and fungi colonized the land, and were soon followed by arthropods and other animals. Insects were particularly successful and even today make up the majority of animal species with the beetles being he largest group. Amphibians first appeared around 300 million years ago, followed by early amniotes, then mammals around 200 million years ago and birds around 100 million years ago (both from “reptile” like lineages). However, despite the evolution of these large animals, smaller organisms similar to the types that evolved early in this process continue to dominate the Earth to this day. This dominance includes both biomass and specie variation with the dominance being the prokaryotes.
Social and cultural responses:
In the 19th century, particularly after the publication of On the Origin of Species in 1859, the idea that life had evolved and not created was an active source of academic debate centred on the philosophical, social and religious implications of evolution. Back as far as Anaximander; 500 BC, who was the author of the first surviving lines of Western philosophy. He speculated and argued about “the Boundless” as the origin of all that is. A basic theory that all life commenced from a single or few individuals. On the other side of the world Zhuangzi (Zhuang Zhou) held similar views. Along with Charles Darwin and Alfred Russel were heavily ridiculed by the religions and the ruling classes of the day. Nowadays, the fact that organisms evolve is uncontested in the scientific literature and the modern evolutionary synthesis is widely accepted by scientists worldwide. Despite the overwhelming evidence and historic records evolution remains a contentious concept for theists.

Darwinism is symbolized by the caricatures of an Apes body with Darwin’s head. It was probably originally drawn as derogatory scepticism but has eventually been accepted and adopted as factual science of evolution.
While various religions and denominations have reconciled their beliefs with evolution through concepts such as theistic evolution, there are creationists who staunchly believe that evolution is contradicted by their creation myths and dispute every aspect of evolution without raising a single fact.
This was clearly demonstrated by responses to the publication of Vestiges of the Natural History of Creation in 1844, the most controversial aspect of evolutionary biology is the implication of human evolution that human mental and moral faculties, which had been thought purely spiritual, are not distinctly separated from those of other animals. In some countries; notably the United States, Germany and Holland, these tensions between science and religion have fueled the current creation evolution controversy. Religious conflict focusing on politics, public education cannot dispute the fact that even evolutionists maintain morality and spiritual awareness. The main antagonists to evolution are the various religious orders that have a lot to lose financially.
“Social Darwinism” is a derogatory term perpetrated and distributed by the Whig philosopher Herbert Spencer. It selectively recollects the ideas about “survival of the fittest” distorting the facts to justify commerce thugery and exploitation by the wealthy ruling class against the working class. It further degenerates human societies as a whole of the fittest should rule; protected by wealth laws, that led to claims that social inequality, sexism, racism, and imperialism were justified. However, these ideas contradict Darwin and Wallace’s own views by both contemporary scientists and philosophers who consider them to be unable to be substantiated.
Applications:
Evolutionary biology, and in particular the understanding of how organisms evolve through natural selection, is an area of science with many practical applications. A major technological application of evolution is artificial selection, which is the intentional selection of certain traits in a population of organisms. Humans have used artificial selection for thousands of years in the domestication of plants and animals. More recently, such selection has become a vital part of genetic engineering, with selectable markers such as antibiotic resistance genes being used to manipulate DNA in molecular biology. It is also possible to use repeated rounds of mutation and selection to evolve proteins with particular properties, such as modified enzymes or new antibodies, in a process called directed evolution.
The changes that have occurred during an organism’s evolution can reveal the genes needed to construct parts of the body, genes which may be involved in human genetic disorders. The Mexican tetra is an albino cavefish that lost its eyesight during evolution. Breeding together different populations of this blind fish produced some offspring with functional eyes, since different mutations had occurred in the isolated populations that had evolved in different caves. This helped identify genes required for vision and pigmentation, such as crystallins and the melanocortin 1 receptor.
Further comments from Members:
All information is included in good faith and has been thoroughly researched prior to printing. The website or the author does not warrant or guarantee the accuracy of any information on these pages, nor does the website or the author accept any responsibility for any loss arising from the use of the information found within. The views and opinions are strictly those of the author or those members who chose to actively participate in the contents herein.
“Hi reader, it seems you use The Bible of Botany a lot. That’s great as we have great pleasure in bringing it to you! It’s a little awkward for us to ask, but our first aim is to purchase land approximately 1,600 hectares to link several parcels of N.P. into one at The Pinnacles NSW Australia, but we need your help. We’re not salespeople. We’re amateur botanists who have dedicated over 30 years to saving the environment in a practical way. We depend on donations to reach our goal. If you donate just $5, the price of your coffee this Sunday, We can help to keep the planet alive in a real way and continue to bring you regular updates and features on Australian plants all in one Botanical Bible. Any support is greatly appreciated. Thank you.”
In the spirit of reconciliation we acknowledge the Bundjalung, Gumbaynggirr and Yaegl and all aboriginal nations throughout Australia and their connections to land, sea and community. We pay our respect to their Elders past, present and future for the pleasures we have gained.